


ਅਰਨਸਟ-ਜਾਨ ਐਗਰਸ1,2, Ate van der Burgt1, Sjaak AW van Heusden2, Michiel E. de Vries@ 1, ਰਿਚਰਡ ਜੀ.ਐਫ. ਵਿਸੇr©2, ਕ੍ਰਿਸ਼ਚੀਅਨ ਡਬਲਯੂ ਬੀ ਬੈਚਮ©2и & ਪਿਮ ਲਿੰਡਹਾਉਟ1
ਆਲੂ ਵਿੱਚ ਜੈਨੇਟਿਕ ਲਾਭ ਕਾਸ਼ਤ ਕੀਤੇ ਆਲੂ ਦੇ ਹੇਟਰੋਜ਼ਾਈਗਸ ਟੈਟਰਾਪਲੋਇਡ ਜੀਨੋਮ ਦੁਆਰਾ ਰੁਕਾਵਟ ਪਾਉਂਦਾ ਹੈ। ਆਲੂ ਨੂੰ ਇੱਕ ਡਿਪਲੋਇਡ ਇਨਬ੍ਰੇਡ-ਲਾਈਨ ਅਧਾਰਤ F1-ਹਾਈਬ੍ਰਿਡ ਫਸਲ ਵਿੱਚ ਤਬਦੀਲ ਕਰਨਾ ਜੈਨੇਟਿਕ ਲਾਭ ਵਧਾਉਣ ਲਈ ਇੱਕ ਵਧੀਆ ਰਸਤਾ ਪ੍ਰਦਾਨ ਕਰਦਾ ਹੈ। ਡਿਪਲੋਇਡ ਆਲੂ ਜਰਮਪਲਾਜ਼ਮ ਵਿੱਚ ਇੱਕ ਪ੍ਰਭਾਵੀ ਐਸ-ਲੋਕਸ ਇਨਿਹਿਬਟਰ (Sli) ਜੀਨ ਦੀ ਸ਼ੁਰੂਆਤ ਸਵੈ-ਉਪਜਾਊ ਬੀਜਾਂ ਦੇ ਕੁਸ਼ਲ ਉਤਪਾਦਨ ਅਤੇ ਇਸ ਤਰ੍ਹਾਂ ਆਲੂਆਂ ਦੀਆਂ ਇਨਬ੍ਰੇਡ ਲਾਈਨਾਂ ਦੇ ਵਿਕਾਸ ਦੀ ਆਗਿਆ ਦਿੰਦੀ ਹੈ। ਸਲੀ ਲੋਕਸ ਦੀ ਬਣਤਰ ਅਤੇ ਕਾਰਜ ਬਾਰੇ ਬਹੁਤ ਘੱਟ ਜਾਣਿਆ ਜਾਂਦਾ ਹੈ। ਇੱਥੇ ਅਸੀਂ ਕ੍ਰੋਮੋਸੋਮ 12.6 'ਤੇ 12 kb ਅੰਤਰਾਲ 'ਤੇ Sli ਦੀ ਮੈਪਿੰਗ ਨੂੰ ਮੁੜ-ਸੰਯੋਗ ਸਕਰੀਨ ਪਹੁੰਚ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਵਰਣਨ ਕਰਦੇ ਹਾਂ। ਇਸ ਅੰਤਰਾਲ ਵਿੱਚ ਮੌਜੂਦ ਦੋ ਉਮੀਦਵਾਰ ਜੀਨਾਂ ਵਿੱਚੋਂ ਇੱਕ ਇੱਕ ਵਿਲੱਖਣ ਕ੍ਰਮ ਦਿਖਾਉਂਦਾ ਹੈ ਜੋ ਵਿਸ਼ੇਸ਼ ਤੌਰ 'ਤੇ ਸਵੈ-ਅਨੁਕੂਲ ਲਾਈਨਾਂ ਵਿੱਚ ਮੌਜੂਦ ਹੁੰਦਾ ਹੈ। ਅਸੀਂ ਇੱਕ ਸਮੀਕਰਨ ਵੈਕਟਰ ਦਾ ਵਰਣਨ ਕਰਦੇ ਹਾਂ ਜੋ ਸਵੈ-ਅਸੰਗਤ ਨੂੰ ਬਦਲਦਾ ਹੈ ਜੀਨੋਟਾਈਪ ਸਵੈ-ਅਨੁਕੂਲ ਅਤੇ ਇੱਕ CRISPR-Cas9 ਵੈਕਟਰ ਵਿੱਚ ਜੋ SC ਜੀਨੋਟਾਈਪਾਂ ਨੂੰ SI ਵਿੱਚ ਬਦਲਦਾ ਹੈ। ਸਲੀ ਜੀਨ ਇੱਕ ਐਫ-ਬਾਕਸ ਪ੍ਰੋਟੀਨ ਨੂੰ ਏਨਕੋਡ ਕਰਦਾ ਹੈ ਜੋ ਵਿਸ਼ੇਸ਼ ਤੌਰ 'ਤੇ ਸਵੈ-ਅਨੁਕੂਲ ਪੌਦਿਆਂ ਦੇ ਪਰਾਗ ਵਿੱਚ ਪ੍ਰਗਟ ਹੁੰਦਾ ਹੈ। ਉਸ ਜੀਨ ਦੇ ਪ੍ਰਮੋਟਰ ਵਿੱਚ ਇੱਕ 533 bp ਸੰਮਿਲਨ ਫੰਕਸ਼ਨ ਮਿਊਟੇਸ਼ਨ ਦੇ ਲਾਭ ਵੱਲ ਲੈ ਜਾਂਦਾ ਹੈ, ਜੋ ਸਵੈ-ਪਰਾਗ ਅਸਵੀਕਾਰਨ ਨੂੰ ਦੂਰ ਕਰਦਾ ਹੈ।
ਆਲੂ ਦੁਨੀਆ ਦੀ ਸਭ ਤੋਂ ਮਹੱਤਵਪੂਰਨ ਗੈਰ-ਅਨਾਜ ਭੋਜਨ ਫਸਲ ਹੈ। ਹਾਲਾਂਕਿ, ਜਦੋਂ ਕਿ ਮੱਕੀ, ਚਾਵਲ ਅਤੇ ਕਣਕ ਵਰਗੀਆਂ ਹੋਰ ਖੁਰਾਕੀ ਫਸਲਾਂ ਨੇ ਪ੍ਰਤੀ ਸਾਲ 1% ਦਾ ਅਨੁਵੰਸ਼ਕ ਉਪਜ ਦਰ ਦਿਖਾਇਆ ਹੈ।1, ਆਲੂ ਵਿੱਚ ਜੈਨੇਟਿਕ ਲਾਭ ਬਹੁਤ ਘੱਟ ਰਿਹਾ ਹੈ2. ਵਰਤਮਾਨ ਵਿੱਚ, ਜ਼ਿਆਦਾਤਰ ਵਪਾਰਕ ਤੌਰ 'ਤੇ ਉਗਾਈਆਂ ਜਾਣ ਵਾਲੀਆਂ ਆਲੂਆਂ ਦੀਆਂ ਕਿਸਮਾਂ ਹੇਟਰੋਜ਼ਾਈਗਸ ਆਟੋਟੇਟ੍ਰੈਪਲੋਇਡ ਮਾਤਾ-ਪਿਤਾ ਦੇ ਵਿਚਕਾਰਲੇ ਕਰਾਸ ਤੋਂ ਪ੍ਰਾਪਤ ਹੁੰਦੀਆਂ ਹਨ। ਇਸ ਪ੍ਰਜਨਨ ਪ੍ਰਣਾਲੀ ਵਿੱਚ, ਉਨ੍ਹਾਂ ਦੁਰਲੱਭ ਵਿਅਕਤੀਆਂ ਦੀ ਪਛਾਣ ਕਰਨ ਲਈ ਹਰ ਇੱਕ ਪ੍ਰਜਨਨ ਪੀੜ੍ਹੀ ਵਿੱਚ ਸੈਂਕੜੇ ਹਜ਼ਾਰਾਂ ਬੂਟੇ ਤਿਆਰ ਕੀਤੇ ਜਾਂਦੇ ਹਨ ਅਤੇ ਉਹਨਾਂ ਦੀ ਜਾਂਚ ਕੀਤੀ ਜਾਂਦੀ ਹੈ ਤਾਂ ਜੋ ਸੰਤਾਨ ਵਿੱਚ ਵੱਖੋ-ਵੱਖਰੇ ਗੁਣਾਂ ਲਈ ਸਵੀਕਾਰਯੋਗ ਵਿਸ਼ੇਸ਼ਤਾਵਾਂ ਹੋਣ। ਜਿਵੇਂ ਕਿ ਇੱਥੇ ਕੁਝ ਪੰਜਾਹ ਗੁਣ ਹਨ ਜੋ ਇੱਕ ਵਪਾਰਕ ਆਲੂ ਦੀ ਕਾਸ਼ਤ ਦੇ ਮੁੱਲ ਲਈ ਢੁਕਵੇਂ ਹਨ, ਪਰੰਪਰਾਗਤ ਆਲੂ ਪ੍ਰਜਨਨ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਇਹਨਾਂ ਗੁਣਾਂ ਨੂੰ ਨਿਯੰਤਰਿਤ ਕਰਨ ਵਾਲੇ ਸਭ ਤੋਂ ਵਧੀਆ ਐਲੀਲਾਂ ਨੂੰ ਜੋੜਨ ਦਾ ਮੌਕਾ ਬਹੁਤ ਘੱਟ ਹੈ। ਇਸ ਤੋਂ ਇਲਾਵਾ, ਬੈਕਕਰੌਸਿੰਗ ਸਕੀਮਾਂ ਰਾਹੀਂ ਜੈਨੇਟਿਕ ਅਖੰਡਤਾ ਨੂੰ ਕਾਇਮ ਰੱਖਦੇ ਹੋਏ ਕੁਲੀਨ ਕਿਸਮਾਂ ਵਿੱਚ ਨਵੇਂ ਗੁਣਾਂ ਦੀ ਨਿਸ਼ਾਨਦੇਹੀ ਕੀਤੀ ਜਾਣ-ਪਛਾਣ ਸਮਲਿੰਗੀ ਮਾਪਿਆਂ ਦੀਆਂ ਲਾਈਨਾਂ ਤੋਂ ਬਿਨਾਂ ਅਸੰਭਵ ਹੈ। ਇਨ੍ਹਾਂ ਸਮੱਸਿਆਵਾਂ ਨੂੰ ਦੂਰ ਕਰਨ ਲਈ, ਕਈ ਸਮੂਹਾਂ ਨੇ ਇਨਬ੍ਰੇਡ-ਲਾਈਨ-ਅਧਾਰਤ ਡਿਪਲੋਇਡ ਆਲੂ ਪ੍ਰਜਨਨ ਪ੍ਰੋਗਰਾਮ ਸ਼ੁਰੂ ਕੀਤੇ ਹਨ।2-5. ਇਹਨਾਂ ਪ੍ਰੋਗਰਾਮਾਂ ਵਿੱਚ, ਪ੍ਰਜਨਨ ਦੌਰਾਨ ਨੁਕਸਾਨਦੇਹ ਐਲੀਲਾਂ ਦੇ ਵਿਰੁੱਧ ਲਗਾਤਾਰ ਚੋਣ ਕਰਕੇ ਅਤੇ ਬੈਕਕਰੌਸਿੰਗ ਸਕੀਮਾਂ ਦੁਆਰਾ ਇਨਬ੍ਰੇਡ ਲਾਈਨਾਂ ਵਿੱਚ ਲਾਭਦਾਇਕ ਐਲੀਲਾਂ ਨੂੰ ਸਟੈਕ ਕਰਕੇ ਮਾਤਾ-ਪਿਤਾ ਦੀਆਂ ਲਾਈਨਾਂ ਦੇ ਵਾਧੇ ਵਾਲੇ ਸੁਧਾਰਾਂ ਦੁਆਰਾ ਜੈਨੇਟਿਕ ਲਾਭ ਪ੍ਰਾਪਤ ਕੀਤੇ ਜਾਂਦੇ ਹਨ।6. ਫਿਰ ਪੇਰੈਂਟਲ ਇਨਬ੍ਰਿਡ ਲਾਈਨਾਂ ਨੂੰ ਹੇਟਰੋਟਿਕ ਐਫ1-ਹਾਈਬ੍ਰਿਡ ਔਲਾਦ ਪੈਦਾ ਕਰਨ ਲਈ ਪਾਰ ਕੀਤਾ ਜਾਂਦਾ ਹੈ।
ਜ਼ਿਆਦਾਤਰ ਡਿਪਲੋਇਡ ਆਲੂ ਜੀਨੋਟਾਈਪਾਂ ਵਿੱਚ, ਇਨਬ੍ਰੀਡਿੰਗ ਇੱਕ ਗੇਮਟੋਫਾਈਟਿਕ ਸਵੈ-ਅਸੰਗਤਤਾ (GSI) ਪ੍ਰਣਾਲੀ ਦੁਆਰਾ ਬੁਰੀ ਤਰ੍ਹਾਂ ਸੀਮਤ ਹੁੰਦੀ ਹੈ ਜੋ ਮਲਟੀ-ਐਲੇਲਿਕ ਐਸ-ਲੋਕਸ ਦੁਆਰਾ ਨਿਯੰਤਰਿਤ ਹੁੰਦੀ ਹੈ। ਇਹ ਐਸ-ਲੋਕਸ ਏਨਕੋਡ ਸਟਾਈਲ ਐਸ-ਆਰਨੇਸ ਨੂੰ ਪ੍ਰਗਟ ਕਰਦਾ ਹੈ ਜੋ ਸ਼ੈਲੀ ਵਿੱਚ ਸਵੈ-ਪਰਾਗ ਟਿਊਬ ਦੇ ਵਿਕਾਸ ਨੂੰ ਰੋਕਦਾ ਹੈ, ਸਵੈ-ਗਰਭਣ ਨੂੰ ਰੋਕਦਾ ਹੈ।7. ਕਰਾਸ-ਪਰਾਗੀਕਰਨ ਦੇ ਦੌਰਾਨ, ਪਰਾਗ-ਪ੍ਰਗਟਿਤ S-ਲੋਕਸ F-ਬਾਕਸ ਪ੍ਰੋਟੀਨ (SLF) S-RNases ਨੂੰ ਪਛਾਣਦੇ ਹਨ ਅਤੇ ਉਹਨਾਂ ਨੂੰ ਪ੍ਰੋਟੀਸੋਮਲ ਡਿਗਰੇਡੇਸ਼ਨ ਮਾਰਗ ਵੱਲ ਨਿਸ਼ਾਨਾ ਬਣਾਉਂਦੇ ਹਨ, ਪਰਾਗ ਟਿਊਬ ਨੂੰ ਅੰਡਕੋਸ਼ਾਂ ਵੱਲ ਵਧਣ ਦੀ ਇਜਾਜ਼ਤ ਦਿੰਦੇ ਹਨ ਜਿੱਥੇ ਗਰੱਭਧਾਰਣ ਹੋ ਸਕਦਾ ਹੈ।8. ਹਰੇਕ S-ਐਲੀਲ ਇੱਕ S-RNase ਅਤੇ ਮਲਟੀਪਲ SLF ਨੂੰ ਵੱਖ-ਵੱਖ ਵਿਸ਼ੇਸ਼ਤਾਵਾਂ ਦੇ ਨਾਲ ਏਨਕੋਡ ਕਰਦਾ ਹੈ, ਜੋ ਇੱਕੋ ਐਲੀਲ 'ਤੇ ਮੌਜੂਦ S-RNase ਨੂੰ ਛੱਡ ਕੇ ਸਾਰੇ S-RNase ਨੂੰ ਪਛਾਣ ਸਕਦੇ ਹਨ।9.
ਹਾਲਾਂਕਿ ਜ਼ਿਆਦਾਤਰ ਡਿਪਲੋਇਡ ਆਲੂ ਲਾਈਨਾਂ ਸਵੈ-ਅਨੁਕੂਲ (SI), ਸਵੈ-ਅਨੁਕੂਲ ਡਿਪਲੋਇਡ ਆਲੂ ਲਾਈਨਾਂ ਮੌਜੂਦ ਹਨ ਅਤੇ ਡਿਪਲੋਇਡ ਆਲੂ ਪ੍ਰਜਨਨ ਪ੍ਰੋਗਰਾਮਾਂ ਵਿੱਚ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਨੂੰ ਪੇਸ਼ ਕਰਨ ਲਈ ਵਰਤੀਆਂ ਜਾ ਸਕਦੀਆਂ ਹਨ।10-12. ਹੋਸਾਕਾ ਅਤੇ ਹੈਨੇਮਨ ਨੇ ਇੱਕ ਪ੍ਰਭਾਵਸ਼ਾਲੀ ਐਸ-ਲੋਕਸ ਇਨਿਹਿਬਟਰ ਨੂੰ ਮੈਪ ਕੀਤਾ (Sli) ਏ ਤੋਂ ਜੀਨ ਸੋਲਨਮ ਚੈਕੋਏਂਸ ਕ੍ਰੋਮੋਸੋਮ 12 ਦੇ ਦੂਰ ਦੇ ਸਿਰੇ 'ਤੇ ਪਹੁੰਚਣਾ ਅਤੇ ਇਸਦੀ ਵਰਤੋਂ ਆਲੂ ਦੀਆਂ ਇਨਬ੍ਰੇਡ ਲਾਈਨਾਂ ਬਣਾਉਣ ਲਈ ਕੀਤੀ।13'14. ਆਪਣੇ ਨਤੀਜਿਆਂ ਦੇ ਆਧਾਰ 'ਤੇ, ਹੋਸਾਕਾ ਅਤੇ ਹੈਨੇਮੈਨ ਨੇ ਸੁਝਾਅ ਦਿੱਤਾ ਕਿ ਸਲੀ ਸਪੋਰੋਫਾਈਟਿਕ ਐਕਸ਼ਨ ਵਾਲਾ ਇੱਕ ਪਰਾਗ-ਪ੍ਰਗਟ ਕੀਤਾ ਜੀਨ ਹੈ ਅਤੇ ਇਸਦੇ ਲਈ ਸਮਰੂਪਤਾ ਹੈ ਸਲੀ ਹੋਮੋਜ਼ਾਈਗਸ ਤੋਂ ਘਾਤਕ ਹੈ ਸਲੀਸਲੀ ਐਸ ਦੀ F8 ਆਬਾਦੀ ਵਿੱਚ ਜੀਨੋਟਾਈਪ ਗੈਰਹਾਜ਼ਰ ਸਨ। chacoense. ਅਸੀਂ ਇਹਨਾਂ ਵਿੱਚੋਂ ਇੱਕ ਐਸ. chacoense (DS) - ਸਵੈ-ਅਨੁਕੂਲਤਾ ਨੂੰ S ਵਿੱਚ ਪੇਸ਼ ਕਰਨ ਲਈ ਉਤਪੰਨ ਇਨਬ੍ਰੇਡ ਲਾਈਨਾਂ। ਕੰਦ ਪਿਛੋਕੜ ਇੱਥੇ, ਅਸੀਂ ਡਿਪਲੋਇਡ ਆਲੂ ਵਿੱਚ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਦੇ ਜੀਵ ਵਿਗਿਆਨ ਵਿੱਚ ਹੋਰ ਸਮਝ ਪ੍ਰਾਪਤ ਕਰਨ ਲਈ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਦੇ ਕਾਰਕ ਜੀਨ ਦੀ ਪਛਾਣ ਦਾ ਵਰਣਨ ਕਰਦੇ ਹਾਂ।
ਨਤੀਜੇ ਅਤੇ ਚਰਚਾ
ਦੇ ਵਿਚਕਾਰ ਇੱਕ ਕਰਾਸ ਤੱਕ ਲਿਆ ਇੱਕ F2 ਆਬਾਦੀ ਵਿੱਚ ਸਲੀ ਦਾਨੀ (ਨਿਯੁਕਤ DS) ਅਤੇ ਇੱਕ ਡਿਪਲੋਇਡ ਐਸ. ਕੰਦ (D2) ਅਸੀਂ ਕ੍ਰੋਮੋਸੋਮ 2 'ਤੇ ਸਵੈ-ਬੇਰੀ ਸੈੱਟ ਲਈ ਇੱਕ ਮਾਮੂਲੀ ਪ੍ਰਭਾਵ QTL ਦੇਖਿਆ, ਪਰ ਬਾਅਦ ਵਿੱਚ ਮੁੜ ਸੰਜੋਗ ਸਕ੍ਰੀਨਿੰਗ ਸਫਲ ਨਹੀਂ ਸੀ। ਅਸੀਂ ਦੇਖਿਆ ਹੈ ਕਿ ਕਈ F2 ਆਬਾਦੀਆਂ ਨੇ ਕ੍ਰੋਮੋਸੋਮ 12 ਦੀ ਲੰਮੀ ਬਾਂਹ ਦੇ ਆਲੇ-ਦੁਆਲੇ ਬਹੁਤ ਜ਼ਿਆਦਾ ਤਿੱਖਾਪਨ ਦਿਖਾਇਆ ਹੈ, ਗੈਰ-ਡੀਐਸ ਹੈਪਲੋਟਾਈਪ ਲਈ ਸਮਰੂਪਤਾ ਪੂਰੀ ਤਰ੍ਹਾਂ ਗੈਰਹਾਜ਼ਰ ਹੈ।
ਇਸ skewness ਅਤੇ ਦੀ ਮੈਪਿੰਗ ਦੇ ਆਧਾਰ 'ਤੇ ਸਲੀ ਕ੍ਰੋਮੋਸੋਮ 12 'ਤੇ, ਅਸੀਂ ਇਹ ਅਨੁਮਾਨ ਲਗਾਇਆ ਹੈ ਸਲੀ ਗੇਮਟੋਫਾਈਟਿਕ ਤੌਰ 'ਤੇ ਪ੍ਰਗਟ ਕੀਤਾ ਗਿਆ ਹੈ, ਮਤਲਬ ਕਿ ਇੱਕ ਪੌਦੇ ਦੇ ਸਵੈ-ਪਰਾਗੀਕਰਨ ਵਿੱਚ ਹੈਟਰੋਜ਼ਾਈਗਸ ਲਈ ਸਲੀ (Sli/sli), ਸਿਰਫ਼ ਪਰਾਗ ਜਿਸ ਵਿੱਚ ਪ੍ਰਬਲ ਹੁੰਦਾ ਹੈ ਸਲੀ ਐਲੀਲ ਸਵੈ-ਗਰਭਕਰਨ ਵਿੱਚ ਹਿੱਸਾ ਲੈ ਸਕਦਾ ਹੈ। ਇਸ ਪਰਿਕਲਪਨਾ ਨੂੰ ਪਰਖਣ ਲਈ, ਅਸੀਂ ਇੱਕ ਜੋਰਦਾਰ ਅਤੇ ਉੱਚ ਸਵੈ-ਉਪਜਾਊ ਰੇਖਾ (16HP1-66) ਨੂੰ ਇੱਕ ਜੋਰਦਾਰ ਸਵੈ-ਅਨੁਕੂਲ ਲਾਈਨ (D16) ਤੱਕ ਪਾਰ ਕੀਤਾ, ਪੂਰੇ-ਜੀਨੋਮ ਦੋਵਾਂ ਨੂੰ ਕ੍ਰਮਬੱਧ ਕੀਤਾ, ਅਤੇ ਨਤੀਜੇ ਵਜੋਂ F1 ਆਬਾਦੀ ਦਾ ਵਿਸ਼ਲੇਸ਼ਣ ਕੀਤਾ (17SC11, n = 251, ਚਿੱਤਰ. 1a, b) ਕਿਉਂਕਿ ਇਹ F1 ਆਬਾਦੀ ਬਹੁਤ ਸਾਰੇ ਸਥਾਨਾਂ ਲਈ ਬਹੁਤ ਜ਼ਿਆਦਾ ਬਹੁਰੂਪੀ ਹੈ, ਅਸੀਂ ਫੀਨੋਟਾਈਪਾਂ ਦੀ ਇੱਕ ਵਿਸ਼ਾਲ ਸ਼੍ਰੇਣੀ ਦੇਖੀ, ਜਿਸ ਵਿੱਚ ਉਹ ਵੀ ਸ਼ਾਮਲ ਹਨ ਜੋ ਉਪਜਾਊ ਸ਼ਕਤੀ ਨਾਲ ਸਬੰਧਤ ਹਨ। ਇਸਲਈ, ਅਸੀਂ ਇੱਕ ਬਹੁਤ ਹੀ ਸਖ਼ਤ ਅਤੇ ਸਖ਼ਤ ਫੀਨੋਟਾਈਪਿੰਗ ਪ੍ਰੋਟੋਕੋਲ ਲਾਗੂ ਕੀਤਾ ਹੈ ਜਿਸ ਵਿੱਚ ਬੇਰੀ ਅਤੇ ਬੀਜਾਂ ਦੇ ਸੈਟ ਅਤੇ ਕ੍ਰਾਸ ਅਤੇ ਸਵੈ-ਪਰਾਗੀਕਰਨ ਦੇ ਨਾਲ-ਨਾਲ ਸਟਾਈਲ ਵਿੱਚ ਪਰਾਗ ਟਿਊਬ ਦੇ ਵਾਧੇ ਦੀ ਕਲਪਨਾ ਸ਼ਾਮਲ ਹੈ ਤਾਂ ਜੋ ਅਨੁਕੂਲਤਾ ਫੀਨੋਟਾਈਪ ਨੂੰ ਉਲਝਾਉਣ ਵਾਲੇ ਨਸਬੰਦੀ ਦੇ ਮੁੱਦਿਆਂ ਤੋਂ ਬਚਿਆ ਜਾ ਸਕੇ। ਜਿਹੜੇ ਪੌਦੇ ਇੱਕ ਤੋਂ ਵੱਧ ਸੈਲਫ-ਬੇਰੀ ਸੈਟ ਕਰਦੇ ਹਨ ਉਹਨਾਂ ਨੂੰ SC ਮੰਨਿਆ ਜਾਂਦਾ ਹੈ, ਜਦੋਂ ਕਿ ਪੌਦੇ ਜੋ ਘੱਟੋ-ਘੱਟ 10 ਸਵੈ-ਪਰਾਗਣ ਤੋਂ ਬਾਅਦ ਸਵੈ-ਬੇਰੀਆਂ ਨੂੰ ਸੈੱਟ ਨਹੀਂ ਕਰਦੇ, ਸ਼ੈਲੀ ਵਿੱਚ ਸਵੈ-ਪਰਾਗ ਟਿਊਬ ਦੇ ਵਾਧੇ ਨੂੰ ਦਰਸਾਉਂਦੇ ਹਨ, ਅਤੇ ਬਲਕ ਨਾਲ ਪਰਾਗਿਤ ਕਰਨ ਤੋਂ ਬਾਅਦ ਕਰਾਸ-ਬੇਰੀ ਸੈੱਟ ਕਰਦੇ ਹਨ। ਪਰਾਗ ਨੂੰ SI ਮੰਨਿਆ ਜਾਂਦਾ ਹੈ। ਨਤੀਜੇ ਵਜੋਂ, ਆਬਾਦੀ ਦੇ ਇੱਕ ਮਹੱਤਵਪੂਰਨ ਹਿੱਸੇ ਨੂੰ ਜੈਨੇਟਿਕ ਵਿਸ਼ਲੇਸ਼ਣਾਂ ਤੋਂ ਬਾਹਰ ਰੱਖਿਆ ਗਿਆ ਸੀ ਕਿਉਂਕਿ ਅਨੁਕੂਲਤਾ ਫੀਨੋਟਾਈਪ ਦਾ ਅਸਪਸ਼ਟ ਤੌਰ 'ਤੇ ਮੁਲਾਂਕਣ ਕਰਨ ਦੀਆਂ ਲੋੜਾਂ ਪੂਰੀਆਂ ਨਹੀਂ ਕੀਤੀਆਂ ਗਈਆਂ ਸਨ। ਫਿਰ ਵੀ, ਬਹੁਗਿਣਤੀ ਆਬਾਦੀ 17SC11 ਸੰਤਾਨ ਦੀ ਅਨੁਕੂਲਤਾ ਸਥਿਤੀ ਦਾ ਮੁਲਾਂਕਣ ਕੀਤਾ ਜਾ ਸਕਦਾ ਹੈ ਅਤੇ ਇਸਨੂੰ ਸਵੈ-ਅਨੁਕੂਲਤਾ (ਪੂਰਕ ਡੇਟਾ 1) ਲਈ ਵੱਖ ਕਰਨ ਲਈ ਦਿਖਾਇਆ ਗਿਆ ਸੀ। ਜਿਵੇਂ ਕਿ ਸਵੈ-ਅਨੁਕੂਲਤਾ 16HP1-66 ਤੋਂ ਉਤਪੰਨ ਹੋਈ ਹੈ, ਅਸੀਂ ਕ੍ਰੋਮੋਸੋਮਸ 2 ਅਤੇ 12 'ਤੇ SNPs ਨੂੰ ਨਿਸ਼ਾਨਾ ਬਣਾਉਣ ਵਾਲੇ KASP ਮਾਰਕਰਾਂ ਨੂੰ ਡਿਜ਼ਾਈਨ ਕਰਨ ਲਈ ਇਸ ਜੀਨੋਟਾਇਪ ਦੇ ਪੂਰੇ ਜੀਨੋਮ ਕ੍ਰਮ ਦੀ ਵਰਤੋਂ ਕੀਤੀ ਹੈ ਜੋ ਕਿ 16HP1-66 ਵਿੱਚ ਵਿਭਿੰਨ ਹਨ ਪਰ ਡੀ16 ਦੀ ਮੈਪਿੰਗ ਵਿੱਚ ਸਮਰੂਪ ਹਨ। ਸਲੀ ਜਣੇਪਾ ਮੇਓਸਿਸ ਵਿੱਚ. ਅਸੀਂ ਇੱਕ ਜੈਨੇਟਿਕ ਨਕਸ਼ਾ ਬਣਾਇਆ, QTL ਵਿਸ਼ਲੇਸ਼ਣ ਕੀਤਾ, ਅਤੇ ਕ੍ਰੋਮੋਸੋਮ 75.72 (ਚਿੱਤਰ 12) ਦੀ ਲੰਬੀ ਬਾਂਹ 'ਤੇ ਇੱਕ ਬਹੁਤ ਮਹੱਤਵਪੂਰਨ QTL (LOD = XNUMX) ਪਾਇਆ। 1b), ਹੋਸਾਕਾ ਅਤੇ ਹੈਨੇਮੈਨ ਦੇ ਨਤੀਜਿਆਂ ਦੀ ਪੁਸ਼ਟੀ ਕਰਦੇ ਹੋਏ।
ਇੱਕ ਵੱਖਰੇ ਜੈਨੇਟਿਕ ਪਿਛੋਕੜ ਵਿੱਚ QTL ਦੀ ਪੁਸ਼ਟੀ ਕਰਨ ਲਈ, ਅਸੀਂ SI ਜੀਨੋਟਾਈਪ D14 ਦੇ ਨਾਲ ਸੋਲਿੰਟਾ ਪ੍ਰਜਨਨ ਪ੍ਰੋਗਰਾਮ ਤੋਂ ਪ੍ਰਾਪਤ ਇੱਕ ਹੋਰ ਉੱਚ ਸਵੈ-ਉਪਜਾਊ ਜੀਨੋਟਾਈਪ ਨੂੰ ਪਾਰ ਕੀਤਾ ਅਤੇ ਨਤੀਜੇ ਵਜੋਂ F1 ਆਬਾਦੀ (17SC25, ਸਪਲੀਮੈਂਟਰੀ ਡੇਟਾ 1) ਦਾ ਵਿਸ਼ਲੇਸ਼ਣ ਕੀਤਾ। ਆਬਾਦੀ 32SC17 ਦੇ 25 ਵਿਅਕਤੀਆਂ ਵਿੱਚੋਂ, ਸਾਨੂੰ ਕੋਈ SI ਵਿਅਕਤੀ ਨਹੀਂ ਮਿਲਿਆ। ਇੱਕ ਵੱਖਰੀ ਆਬਾਦੀ ਪੈਦਾ ਕਰਨ ਲਈ, ਅਸੀਂ ਸਭ ਤੋਂ ਉਪਜਾਊ ਜੀਨੋਟਾਈਪ ਨੂੰ ਚੁਣਿਆ ਅਤੇ ਇਸਨੂੰ ਦੋ SI ਜੀਨੋਟਾਈਪਾਂ ਤੱਕ ਪਾਰ ਕੀਤਾ ਜੋ ਅਸੀਂ ਆਬਾਦੀ 17SC11 ਵਿੱਚ ਪਛਾਣੇ ਹਨ, ਨਤੀਜੇ ਵਜੋਂ ਆਬਾਦੀ 18SC11 ਅਤੇ 18SC12 (ਪੂਰਕ ਡੇਟਾ 1 ਅਤੇ ਪੂਰਕ ਚਿੱਤਰ 1) ਹੈ। ਜਿਵੇਂ ਕਿ ਉਮੀਦ ਕੀਤੀ ਜਾਂਦੀ ਹੈ, ਆਬਾਦੀ 18SC11 ਅਤੇ 18SC12 ਦੇ ਵਿਸ਼ਲੇਸ਼ਣ ਨੇ ਦਿਖਾਇਆ ਹੈ ਕਿ ਦੋਵੇਂ ਆਬਾਦੀ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਲਈ ਵੱਖ-ਵੱਖ ਹਨ। ਅਸੀਂ ਪੂਰੇ ਜੀਨੋਮ ਕ੍ਰਮ ਲਈ ਮਾਂ (17SC25-8) ਨੂੰ ਜਮ੍ਹਾ ਕੀਤਾ ਅਤੇ ਇਸ ਡੇਟਾ ਦੀ ਵਰਤੋਂ 17SC11 ਦੀ ਆਬਾਦੀ ਲਈ ਵਰਤੀ ਗਈ ਪਹੁੰਚ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਨਵੇਂ KASP ਮਾਰਕਰਾਂ ਨੂੰ ਡਿਜ਼ਾਈਨ ਕਰਨ ਲਈ ਕੀਤੀ, ਪਰ ਇਸ ਵਾਰ ਸਿਰਫ ਕ੍ਰੋਮੋਸੋਮ 12 ਨੂੰ ਨਿਸ਼ਾਨਾ ਬਣਾਇਆ ਗਿਆ। ਬਾਅਦ ਦੇ QTL ਵਿਸ਼ਲੇਸ਼ਣ ਨੇ QTL ਦੀ ਪੁਸ਼ਟੀ ਕੀਤੀ ਜੋ ਸਾਨੂੰ ਆਬਾਦੀ ਵਿੱਚ ਪਾਇਆ ਗਿਆ ਸੀ। ਕ੍ਰਮਵਾਰ 17SC11 ਅਤੇ 33.14SC120.94 ਆਬਾਦੀ ਵਿੱਚ 18 ਅਤੇ 11 ਦੇ LOD ਮੁੱਲਾਂ ਦੇ ਨਾਲ 18SC12 (ਪੂਰਕ ਚਿੱਤਰ 2)।
ਇਹ ਨਿਰਧਾਰਤ ਕਰਨ ਲਈ ਕਿ ਕੀ ਸਲੀ ਅਸਲ ਵਿੱਚ ਗੇਮਟੋਫਾਈਟਿਕ ਤੌਰ ਤੇ ਪ੍ਰਗਟ ਕੀਤਾ ਗਿਆ ਹੈ, ਅਸੀਂ ਇੱਕ F2 ਆਬਾਦੀ ਦਾ ਵਿਸ਼ਲੇਸ਼ਣ ਕੀਤਾ (19SC1, n = 160) ਇੱਕ ਉਪਜਾਊ ਅਤੇ ਜੋਸ਼ਦਾਰ 17SC11 ਵਿਅਕਤੀ ਤੋਂ ਲਿਆ ਗਿਆ ਹੈ। F1 ਦੇ ਮੁਕਾਬਲੇ ਇਸ ਆਬਾਦੀ ਵਿੱਚ ਫੁੱਲ ਅਤੇ ਉਪਜਾਊ ਸ਼ਕਤੀ ਘੱਟ ਗਈ ਸੀ। ਫੀਨੋਟਾਈਪਿਕ ਵਿਸ਼ਲੇਸ਼ਣ ਵਿੱਚ, 160 ਪੌਦਿਆਂ ਵਿੱਚੋਂ, 81 ਪੌਦੇ ਸਵੈ-ਅਨੁਕੂਲ ਸਨ, 78 ਨੂੰ ਮਾੜੇ ਫੁੱਲਾਂ ਜਾਂ ਮਾੜੀ ਉਪਜਾਊ ਸ਼ਕਤੀ ਦੇ ਕਾਰਨ ਨਿਰਧਾਰਤ ਨਹੀਂ (ਐਨਡੀ) ਵਜੋਂ ਸ਼੍ਰੇਣੀਬੱਧ ਕੀਤਾ ਗਿਆ ਸੀ ਅਤੇ ਇੱਕ ਪੌਦੇ ਨੂੰ ਸਵੈ-ਅਨੁਕੂਲ (ਪੂਰਕ ਡੇਟਾ 1) ਦਾ ਦਰਜਾ ਦਿੱਤਾ ਗਿਆ ਸੀ। ਅਸੀਂ ਕ੍ਰੋਮੋਸੋਮ 12 'ਤੇ SNPs ਨੂੰ ਨਿਸ਼ਾਨਾ ਬਣਾਉਣ ਵਾਲੇ KASP ਮਾਰਕਰ ਡਿਜ਼ਾਈਨ ਕੀਤੇ ਹਨ ਜੋ ਮਾਪਿਆਂ 16HP1-66 ਅਤੇ D16 ਵਿੱਚ ਵਿਕਲਪਕ ਐਲੀਲਾਂ ਲਈ ਸਮਰੂਪ ਹਨ। ਪੂਰੇ ਕ੍ਰੋਮੋਸੋਮ ਦੇ ਨਾਲ 12 ਅਲੱਗ-ਥਲੱਗ ਅਨੁਪਾਤ ਸੰਭਾਵਿਤ 1:2:1 ਅਲੱਗ-ਥਲੱਗ ਤੋਂ ਮਹੱਤਵਪੂਰਨ ਤੌਰ 'ਤੇ ਭਟਕ ਜਾਂਦੇ ਹਨ। ਇਸ ਤੋਂ ਇਲਾਵਾ, ਸਵੈ-ਅਨੁਕੂਲਤਾ QTL ਦੇ ਆਲੇ-ਦੁਆਲੇ, ਪੇਰੈਂਟ ਡੀ 16 (ਪੂਰਕ ਚਿੱਤਰ 3) ਦੇ ਹੈਪਲੋਟਾਈਪ ਲਈ ਕੋਈ ਲੋਕੀ ਹੋਮੋਜ਼ਾਈਗਸ ਨਹੀਂ ਹੈ, ਜਿਸ ਦੀ ਬਜਾਏ ਹੇਟਰੋਜ਼ਾਈਗਸ D1/1HP16-16: ਸਮਰੂਪ 1HP66-16 ਲਈ 1:66 ਵਿਭਾਜਨ ਦਿਖਾਉਂਦਾ ਹੈ, ਜੋ ਸੁਝਾਅ ਦਿੰਦਾ ਹੈ ਕਿ ਪਰਾਗ ਦੀ ਘਾਟ ਸਲੀ ਵੱਖ-ਵੱਖ ਵਿਗਾੜ ਦਾ ਕਾਰਨ ਬਣਦਾ ਹੈ. ਇਹ ਇਸ ਪਰਿਕਲਪਨਾ ਦਾ ਸਮਰਥਨ ਕਰਦਾ ਹੈ ਕਿ ਸਿਰਫ ਪਰਾਗ ਹੀ ਪ੍ਰਭਾਵੀ ਹੁੰਦਾ ਹੈ ਸਲੀ ਐਲੀਲ ਸਵੈ-ਰੱਖਣ ਵਿੱਚ ਹਿੱਸਾ ਲੈਂਦੇ ਹਨ। ਇਸ ਤੋਂ ਇਲਾਵਾ, ਇੱਕ ਵਿਅਕਤੀ ਤੋਂ ਇਲਾਵਾ, SI ਅਤੇ SC ਵਿਚਕਾਰ ਅੰਤਰ ਲਈ ਫੀਨੋਟਾਈਪਿੰਗ ਨਿਰਣਾਇਕ ਸੀ, ਇਹ ਦਰਸਾਉਂਦੀ ਹੈ ਕਿ ਵਰਤਿਆ ਗਿਆ ਫੀਨੋਟਾਈਪਿੰਗ ਪ੍ਰੋਟੋਕੋਲ ਮਜ਼ਬੂਤ ਅਤੇ ਲਗਭਗ ਗਲਤੀ-ਮੁਕਤ ਹੈ।
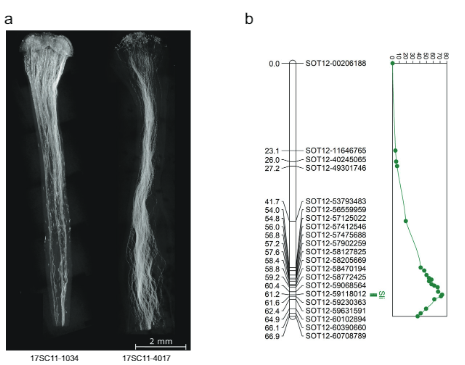
ਜਦਕਿ 628 ਕੇ.ਬੀ SLI ਆਬਾਦੀ 12SC17 ਤੋਂ ਸਲੀ-ਐਲੀਲ ਨੂੰ ਲੈ ਕੇ ਜਾਣ ਵਾਲੇ ਕ੍ਰੋਮੋਸੋਮ 11 'ਤੇ ਅੰਤਰਾਲ ਨੂੰ ਆਬਾਦੀ 169SC18 ਵਿੱਚ 12 KB ਦੇ ਇੱਕ ਛੋਟੇ ਓਵਰਲੈਪਿੰਗ ਅੰਤਰਾਲ ਤੱਕ ਘਟਾ ਦਿੱਤਾ ਗਿਆ ਸੀ, ਇਹ ਅੰਤਰਾਲ ਅਜੇ ਵੀ ਸਲੀ-ਜੀਨ ਦੀ ਪਛਾਣ ਕਰਨ ਲਈ ਬਹੁਤ ਵੱਡੇ ਸਨ। ਇਸ ਲਈ, ਅਸੀਂ ਇੱਕ ਰੀਕੌਂਬੀਨੈਂਟ ਸਕ੍ਰੀਨਿੰਗ ਪਹੁੰਚ ਦੁਆਰਾ S/i- ਰੱਖਣ ਵਾਲੇ ਅੰਤਰਾਲ ਨੂੰ ਘਟਾਉਣ ਦਾ ਟੀਚਾ ਰੱਖਿਆ ਹੈ। ਵਿੱਚ ਪੁਨਰ-ਸੰਯੋਜਨ ਦੇ ਨਾਲ ਪੌਦਿਆਂ ਦੀ ਪਛਾਣ ਕਰਨ ਲਈ ਸਲੀ ਅੰਤਰਾਲ, ਅਸੀਂ 1374 17SC11 ਬੀਜਾਂ ਨੂੰ ਦੋ KASP ਮਾਰਕਰਾਂ ਨਾਲ ਨੇੜਲਾ ਸਰਹੱਦ 'ਤੇ ਅਤੇ ਦੋ ਦੂਰ-ਦੂਰ ਦੀ ਸਰਹੱਦ 'ਤੇ ਜੀਨੋਟਾਈਪ ਕੀਤਾ। ਅਸੀਂ ਦੋ ਬਾਹਰੀ ਮਾਰਕਰਾਂ ਦੇ ਵਿਚਕਾਰ ਪੁਨਰ-ਸੰਯੋਜਨ ਦੇ ਨਾਲ 81 ਬੂਟਿਆਂ ਦੀ ਪਛਾਣ ਕੀਤੀ ਅਤੇ ਉਹਨਾਂ ਨੂੰ ਹੋਰ ਵਧੀਆ ਮੈਪਿੰਗ ਲਈ ਚੁਣਿਆ। ਅਸਪਸ਼ਟ ਫੀਨੋਟਾਈਪਾਂ ਨੂੰ ਪ੍ਰਾਪਤ ਕਰਨ ਲਈ, ਅਸੀਂ ਇਨ੍ਹਾਂ ਜੀਨੋਟਾਈਪਾਂ ਨੂੰ ਬਨਸਪਤੀ ਤੌਰ 'ਤੇ ਪ੍ਰਸਾਰਿਤ ਕੀਤਾ ਅਤੇ ਪ੍ਰਤੀ ਜੀਨੋਟਾਈਪ 'ਤੇ ਘੱਟੋ-ਘੱਟ ਦੋ ਕਲੋਨਾਂ 'ਤੇ ਫੀਨੋਟਾਈਪਿੰਗ ਕੀਤੀ। ਅਸੀਂ ਅੰਤਰਾਲ ਵਿੱਚ ਹੋਰ ਮਾਰਕਰਾਂ ਦੇ ਨਾਲ 81 ਰੀਕੌਂਬੀਨੈਂਟਸ ਨੂੰ ਜੀਨੋਟਾਈਪ ਕੀਤਾ ਅਤੇ ਦੋ ਸੂਚਨਾਤਮਕ ਰੀਕੌਂਬੀਨੈਂਟਸ ਦੀ ਪਛਾਣ ਕੀਤੀ ਜਿਨ੍ਹਾਂ ਨੇ ਅੰਤਰਾਲ ਨੂੰ 27.37 KB ਤੱਕ ਘਟਾ ਦਿੱਤਾ ਜਿਸ ਵਿੱਚ ਪੰਜ ਐਨੋਟੇਟਿਡ ਜੀਨਾਂ (ਪੂਰਕ ਡੇਟਾ 1) ਹਨ।
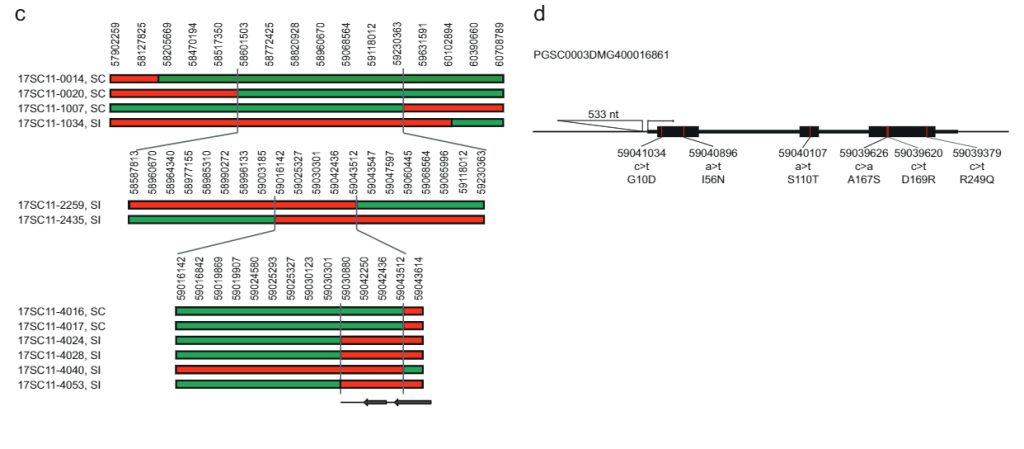
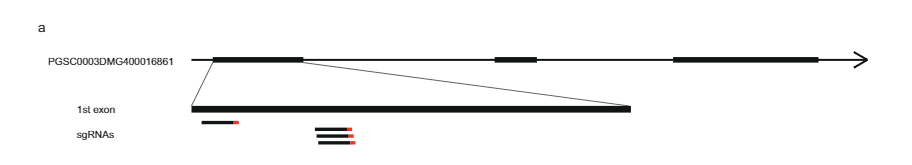
ਅੰਤਰਾਲ ਨੂੰ ਹੋਰ ਘਟਾਉਣ ਲਈ, ਅਸੀਂ ਇਸ 10165 KB ਅੰਤਰਾਲ ਦੇ ਆਲੇ-ਦੁਆਲੇ ਚਾਰ ਮਾਰਕਰਾਂ ਦੇ ਨਾਲ ਉਸੇ ਆਬਾਦੀ ਤੋਂ ਇੱਕ ਹੋਰ 27.37 ਬੂਟਿਆਂ ਦੀ ਜਾਂਚ ਕੀਤੀ ਅਤੇ 12 ਰੀਕੌਂਬੀਨੈਂਟਸ ਦੀ ਪਛਾਣ ਕੀਤੀ। ਇਹਨਾਂ ਨੂੰ ਇਸ ਅੰਤਰਾਲ ਵਿੱਚ 14 ਹੋਰ ਮਾਰਕਰਾਂ ਨਾਲ ਅੱਗੇ ਜੀਨੋਟਾਈਪ ਕੀਤਾ ਗਿਆ ਸੀ ਅਤੇ ਅਸੀਂ ਛੇ ਜਾਣਕਾਰੀ ਭਰਪੂਰ ਰੀਕੌਂਬੀਨੈਂਟਸ ਦੀ ਪਛਾਣ ਕੀਤੀ ਜੋ ਸਪਸ਼ਟ ਅਨੁਕੂਲਤਾ ਫੀਨੋਟਾਈਪ ਦਿਖਾਉਂਦੇ ਹਨ। ਇੱਕ SC ਫੀਨੋਟਾਈਪ ਵਾਲੇ ਦੋ ਰੀਕੌਂਬੀਨੈਂਟਸ ਅਤੇ ਇੱਕ SI ਫੀਨੋਟਾਈਪ ਵਾਲੇ ਇੱਕ ਨੇ 27.37 KB ਅੰਤਰਾਲ ਦੀ ਦੂਰ-ਦੂਰ ਦੀ ਸਰਹੱਦ ਦੀ ਪੁਸ਼ਟੀ ਕੀਤੀ, ਜਦੋਂ ਕਿ ਇੱਕ SI ਫੀਨੋਟਾਈਪ ਵਾਲੇ ਤਿੰਨ ਰੀਕੌਂਬੀਨੈਂਟਾਂ ਨੇ ਇੱਕ ਨਵੀਂ ਪ੍ਰੌਕਸੀਮਲ ਬਾਰਡਰ ਨੂੰ ਪਰਿਭਾਸ਼ਿਤ ਕੀਤਾ ਜੋ ਅੰਤਰਾਲ ਨੂੰ ਸਿਰਫ 12.6 kb ਤੱਕ ਘਟਾ ਦਿੰਦਾ ਹੈ ਜਿਸ ਵਿੱਚ ਦੋ ਜੀਨਾਂ, PGSC0003MG400016861MG0003MG ਅਤੇ ਅੰਜੀਰ. 1c).
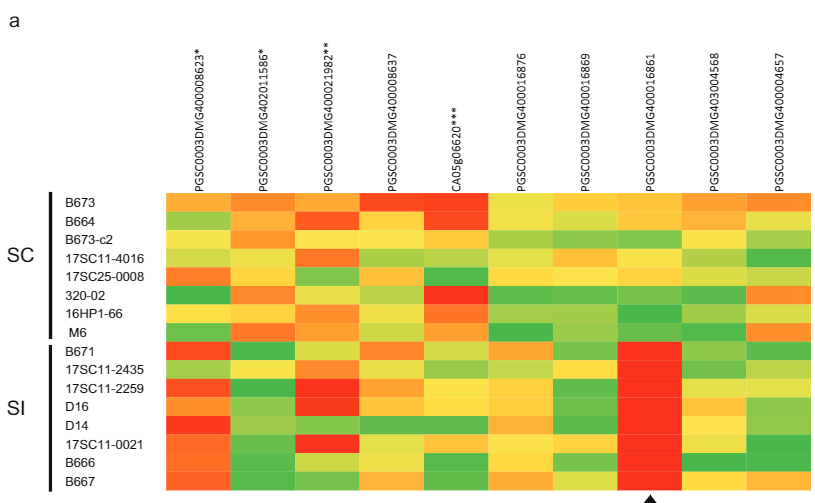
ਉਮੀਦਵਾਰ ਜੀਨ ਦੀ ਪਛਾਣ ਕਰਨ ਲਈ ਜੋ ਸਵੈ-ਅਨੁਕੂਲ ਫੀਨੋਟਾਈਪ ਲਈ ਜ਼ਿੰਮੇਵਾਰ ਹੈ, ਅਸੀਂ ਕਈ ਪੂਰੇ-ਜੀਨੋਮ ਕ੍ਰਮਵਾਰ ਡਿਪਲੋਇਡ ਆਲੂ ਲਾਈਨਾਂ (ਪੂਰਕ ਡੇਟਾ 2) ਵਿੱਚ ਇਹਨਾਂ ਦੋ ਜੀਨਾਂ ਲਈ ਕ੍ਰਮ ਪਰਿਵਰਤਨ ਦਾ ਵਿਸ਼ਲੇਸ਼ਣ ਕੀਤਾ। ਇਹਨਾਂ ਲਾਈਨਾਂ ਦੇ SC/SI ਫੀਨੋਟਾਈਪਾਂ ਨਾਲ ਇਸ ਕ੍ਰਮ ਪਰਿਵਰਤਨ ਦੀ ਤੁਲਨਾ ਕਰਕੇ, ਅਸੀਂ ਸਾਰੇ SC-ਵਿਸ਼ੇਸ਼ SNPs ਅਤੇ INDELS (ਪੂਰਕ ਡੇਟਾ 2) ਦੀ ਪਛਾਣ ਕੀਤੀ ਹੈ। ਅੱਗੇ, ਅਸੀਂ ਹੱਥੀਂ ਸਾਰੇ ਗੈਰ-ਸਮਾਨਯੋਗ SNPs ਦੀ ਪਛਾਣ ਕੀਤੀ ਅਤੇ ਇਹ ਨਿਰਧਾਰਿਤ ਕੀਤਾ ਕਿ ਕੀ ਅਮੀਨੋ ਐਸਿਡ ਦੇ ਬਦਲ ਆਮ ਹਨ ਜਾਂ ਸੋਲਾਨੇਸੀ ਵਿੱਚ ਸਮਾਨ ਪ੍ਰੋਟੀਨ ਲਈ ਵਿਲੱਖਣ ਹਨ। ਉਮੀਦਵਾਰ ਜੀਨ PGSC0003DMG400016861 ਛੇ SC-ਵਿਸ਼ੇਸ਼ ਅਮੀਨੋ ਐਸਿਡ ਬਦਲ ਦਿਖਾਉਂਦਾ ਹੈ ਅਤੇ ਖਾਸ ਤੌਰ 'ਤੇ, ਸਟਾਰਟ ਕੋਡਨ ਤੋਂ —533 bp 'ਤੇ ਸਥਿਤ ਇੱਕ 108 bp ਸੰਮਿਲਨ, ਸੁਝਾਅ ਦਿੰਦਾ ਹੈ ਕਿ SC ਐਲੀਲ ਨੇ SI ਐਲੀਲ ਦੇ ਮੁਕਾਬਲੇ ਸਮੀਕਰਨ ਬਦਲਿਆ ਹੈ। ਇਹਨਾਂ ਜੈਨੇਟਿਕ ਅਧਿਐਨਾਂ ਦੇ ਅਧਾਰ ਤੇ ਅਸੀਂ ਇਹ ਅਨੁਮਾਨ ਲਗਾਇਆ ਹੈ ਕਿ PGSC0003DMG400016861 ਹੈ ਸਲੀ ਜੀਨ.
ਇਸ ਧਾਰਨਾ ਦੀ ਪੁਸ਼ਟੀ ਕਰਨ ਲਈ ਕਿ Sli ਨੂੰ ਪਰਾਗ ਵਿੱਚ ਦਰਸਾਇਆ ਗਿਆ ਹੈ, ਅਸੀਂ ਵਿਟਰੋ ਵਿੱਚ 10 SI ਅਤੇ 10 SC ਆਲੂ ਜੀਨੋਟਾਈਪਾਂ ਤੋਂ ਪਰਾਗ ਨੂੰ ਉਗਾਇਆ ਅਤੇ RNA ਕ੍ਰਮ ਲਈ RNA ਕੱਢਿਆ। ਬਾਕੀ ਬਚੇ ਦੋ ਉਮੀਦਵਾਰ ਜੀਨਾਂ ਵਿੱਚੋਂ, ਸਿਰਫ਼ ਉਮੀਦਵਾਰ ਜੀਨ PGSC0003DMG400016861 ਪ੍ਰਗਟ ਕੀਤਾ ਗਿਆ ਸੀ, ਪਰ ਵਿਸ਼ੇਸ਼ ਤੌਰ 'ਤੇ SC ਜੀਨੋਟਾਈਪਾਂ (ਚਿੱਤਰ. 2a). ਇਸ ਤੋਂ ਇਲਾਵਾ, ਪੋਟੇਟਿਵ ਉਮੀਦਵਾਰ ਸਲੀ ਜੀਨ ਲਈ ਵਿਭਿੰਨ ਪੌਦਿਆਂ ਵਿੱਚ, ਸਿਰਫ ਸਲੀ ਐਲੀਲ ਨੂੰ ਦਰਸਾਇਆ ਗਿਆ ਸੀ। ਦਿਲਚਸਪ ਗੱਲ ਇਹ ਹੈ ਕਿ, ਕ੍ਰੋਮੋਸੋਮ 12 'ਤੇ ਸਲੀ ਲੋਕਸ ਦੇ ਨੇੜੇ ਸਥਿਤ ਹੋਰ ਪਰਾਗ-ਪ੍ਰਗਟ ਕੀਤੇ ਜੀਨਾਂ ਨੇ SC ਅਤੇ SI ਪੌਦਿਆਂ (ਚਿੱਤਰ XNUMX) ਵਿੱਚ ਸਮਾਨ ਪ੍ਰਗਟਾਵਾ ਪੱਧਰ ਦਿਖਾਇਆ। 2a). ਇਸ ਲਈ, ਅਸੀਂ ਸਿੱਟਾ ਕੱਢਿਆ ਹੈ ਕਿ ਕੇਵਲ PGSC0003DMG400016861 ਜੀਨ ਵਿਸ਼ੇਸ਼ ਤੌਰ 'ਤੇ SC ਪੌਦਿਆਂ ਦੀਆਂ ਪਰਾਗ ਟਿਊਬਾਂ ਵਿੱਚ ਪ੍ਰਗਟ ਹੁੰਦਾ ਹੈ।
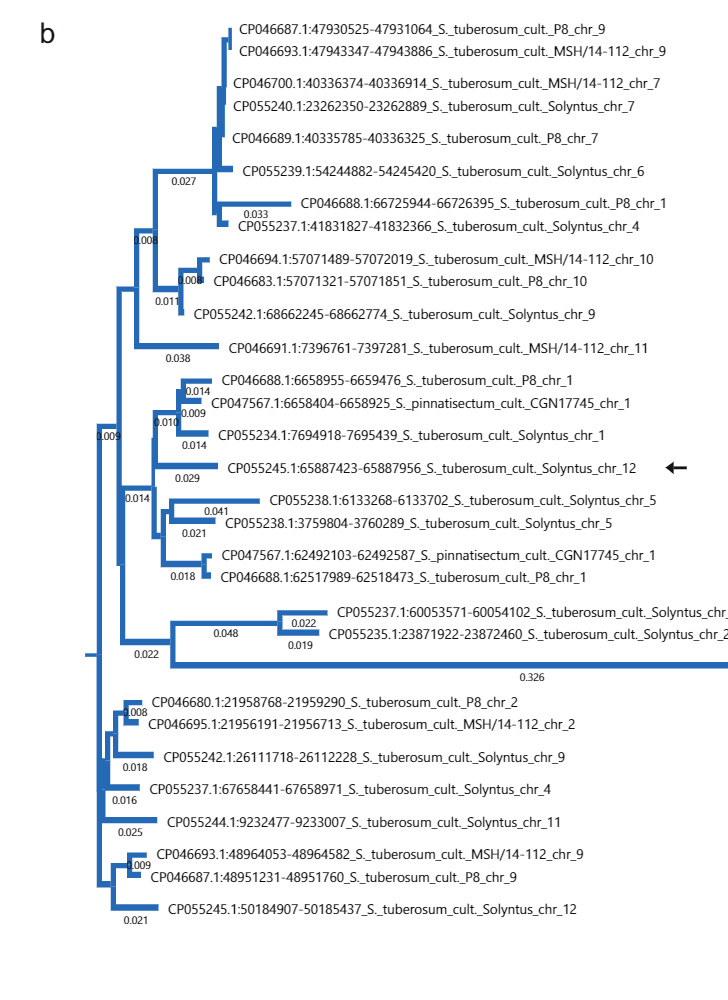
533 bp ਸੰਮਿਲਨ ਦੇ ਮੂਲ ਦੀ ਜਾਂਚ ਕਰਨ ਲਈ, ਅਸੀਂ NCBI 'ਤੇ 533 bp ਕ੍ਰਮ ਦੀ ਇੱਕ BLAST ਖੋਜ ਕੀਤੀ। ਦਿਲਚਸਪ ਗੱਲ ਇਹ ਹੈ ਕਿ, Sli-ਵਿਸ਼ੇਸ਼ ਸੰਮਿਲਨ ਦੇ ਸਮਾਨ ਕ੍ਰਮ ਕ੍ਰਮਬੱਧ S ਵਿੱਚ ਆਮ ਹਨ. ਕੰਦ ਪਹੁੰਚ (ਚਿੱਤਰ. 2b). ਇਸ ਤੋਂ ਇਲਾਵਾ, 533 bp ਸੰਮਿਲਨ ਵਿੱਚ S ਵਿੱਚ ਇੱਕ ਕ੍ਰਮ ਦੀ ਸਮਰੂਪਤਾ ਹੈ. ਪੈਨੇਲੀ ਐਸ ਤੋਂ ਕ੍ਰਮ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ. ਪੈਨੇਲੀ ਇੱਕ BLAST ਪੁੱਛਗਿੱਛ ਦੇ ਰੂਪ ਵਿੱਚ ਸਾਨੂੰ ਐਸ ਵਿੱਚ ਸਮਾਨ ਕ੍ਰਮ ਮਿਲੇ ਹਨ. lycopersicum. ਵਿੱਚ ਕ੍ਰਮਾਂ ਦਾ ਫਾਈਲੋਜੈਨੇਟਿਕ ਵਿਸ਼ਲੇਸ਼ਣ ਐਸ ਟਿosਬਰੋਸਮ, ਐਸ. ਪੈਨੇਲੀ ਅਤੇ S. lycopersicum ਗਰੁੱਪ ਐਸ. ਪੈਨੇਲੀ ਦੇ ਨਾਲ ਮਿਲ ਕੇ ਕ੍ਰਮ S. lycopersicum ਅਤੇ ਇਕ ਐਸ ਟਿosਬਰੋਸਮ ਕ੍ਰਮ, ਇਹ ਸੁਝਾਅ ਦਿੰਦਾ ਹੈ ਕਿ ਇਹ ਇੱਕ ਸਾਂਝੇ ਮੂਲ ਨੂੰ ਸਾਂਝਾ ਕਰਦੇ ਹਨ (ਪੂਰਕ ਚਿੱਤਰ 4a)। ਅਸੀਂ ਇਹ ਅਨੁਮਾਨ ਲਗਾਇਆ ਹੈ ਕਿ ਸੰਮਿਲਨ ਇੱਕ ਟ੍ਰਾਂਸਪੋਸੇਬਲ ਐਲੀਮੈਂਟ (TE) ਤੋਂ ਲਿਆ ਗਿਆ ਹੈ। ਅਸੀਂ 533 bp ਕ੍ਰਮ ਤੋਂ ਇੱਕ ਡਾਟ-ਪਲਾਟ ਗ੍ਰਾਫ਼ ਤਿਆਰ ਕੀਤਾ ਅਤੇ ਦੇਖਿਆ ਕਿ ਕ੍ਰਮ ਵਿੱਚ ਛੋਟੇ ਉਲਟ ਦੁਹਰਾਓ (ਪੂਰਕ ਚਿੱਤਰ 4b) ਸ਼ਾਮਲ ਹਨ। ਅਸੀਂ ਪਲਾਂਟ MITE ਡੇਟਾਬੇਸ ਦੇ ਵਿਰੁੱਧ BLAST ਨੂੰ 533 bp ਸੰਮਿਲਨ ਜਮ੍ਹਾ ਕੀਤਾ, ਨਤੀਜੇ ਵਜੋਂ MITE ਪਰਿਵਾਰ DTA_Sot42 ਤੋਂ ਕਈ ਹਿੱਟ ਹੋਏ ਐਸ ਟਿosਬਰੋਸਮ15ਦੇ ਪ੍ਰਮੋਟਰ ਵਿੱਚ 533 ਬੀਪੀ ਸੰਮਿਲਨ ਨੂੰ ਦਰਸਾਉਂਦਾ ਹੈ ਸਲੀ ਅਸਲ ਵਿੱਚ ਇੱਕ TE (ਪੂਰਕ ਚਿੱਤਰ 4c) ਤੋਂ ਉਤਪੰਨ ਹੁੰਦਾ ਹੈ।
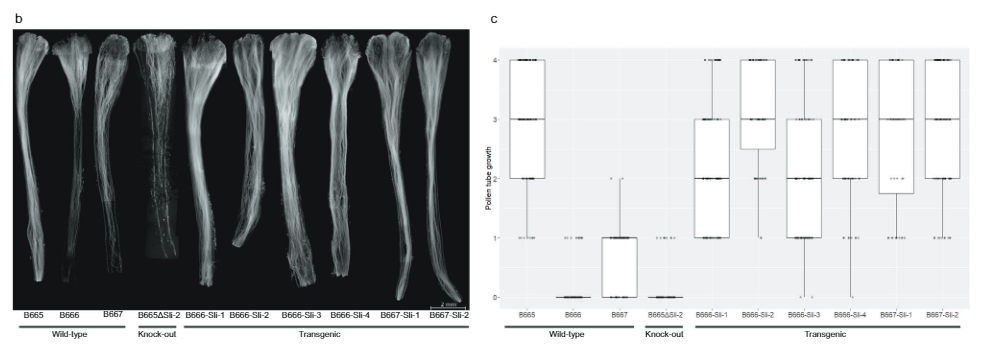
ਹੋਰ ਪੁਸ਼ਟੀ ਕਰਨ ਲਈ ਕਿ PGSC0003DMG400016861 ਅਸਲ ਵਿੱਚ ਹੈ ਸਲੀ, ਅਸੀਂ ਇੱਕ ਸਮੀਕਰਨ ਕੰਸਟਰੱਕਟ ਤਿਆਰ ਕੀਤਾ ਹੈ ਜਿਸ ਵਿੱਚ SC ਐਲੀਲ ਦੇ ਐਕਸੌਨ ਹੁੰਦੇ ਹਨ ਸਲੀ ਇਸਦੇ ਮੂਲ ਪ੍ਰਮੋਟਰ ਅਤੇ ਟਰਮੀਨੇਟਰ ਦੇ ਵਿਚਕਾਰ (ਚਿੱਤਰ. 3a) ਵੈਕਟਰ pBINPLUS (pBINPLUS-Sli) ਵਿੱਚ। ਅਸੀਂ ਇਸ ਨਿਰਮਾਣ ਦੀ ਵਰਤੋਂ ਆਬਾਦੀ 18SC12 ਦੀ ਮੈਪਿੰਗ ਤੋਂ ਦੋ SI ਜੀਨੋਟਾਈਪਾਂ ਨੂੰ ਬਦਲਣ ਲਈ ਕੀਤੀ ਹੈ। ਅਸੀਂ SI ਜੀਨੋਟਾਈਪ B666 ਤੋਂ ਲਏ ਗਏ ਪੰਜ ਸੁਤੰਤਰ ਟ੍ਰਾਂਸਜੇਨਿਕਾਂ ਵਿੱਚੋਂ ਹਰੇਕ ਵਿੱਚੋਂ ਦੋ ਤੋਂ ਛੇ ਕਲੋਨ, ਅਤੇ SI ਜੀਨੋਟਾਈਪ B667 ਤੋਂ ਪ੍ਰਾਪਤ ਤਿੰਨ ਟ੍ਰਾਂਸਜੇਨਿਕਾਂ ਵਿੱਚੋਂ ਦੋ ਤੋਂ ਛੇ ਕਲੋਨ ਫਿਨੋਟਾਈਪ ਕੀਤੇ।
ਛੇ ਸੁਤੰਤਰ ਟ੍ਰਾਂਸਜੇਨਿਕਾਂ ਦੇ ਕਲੋਨ ਸਵੈ-ਪਰਾਗਣ (ਪੂਰਕ ਡੇਟਾ 3) 'ਤੇ ਆਸਾਨੀ ਨਾਲ ਬੇਰੀਆਂ ਨੂੰ ਸੈੱਟ ਕਰਦੇ ਹਨ। ਇਸ ਤੋਂ ਇਲਾਵਾ, ਫਲੋਰੋਸੈਂਸ ਮਾਈਕ੍ਰੋਸਕੋਪੀ ਨੇ ਦਿਖਾਇਆ ਕਿ ਪਰਾਗ ਤੋਂ ਸਲੀ ਟਰਾਂਸਜੇਨਿਕ ਪੌਦੇ ਅਣ-ਪਰਿਵਰਤਿਤ ਨਿਯੰਤਰਣਾਂ ਨਾਲੋਂ ਸਵੈ-ਸ਼ੈਲੀ ਵਿੱਚ ਡੂੰਘੇ ਵਧਦੇ ਹਨ (ਚਿੱਤਰ. 3b, c) ਅਸੀਂ ਵਿੱਚ 442 ਸਵੈ-ਪਰਾਗਣਾਂ ਵਿੱਚ ਪਰਾਗ ਟਿਊਬ ਵਿੱਚ ਵਾਧਾ ਦਰਜ ਕੀਤਾ ਸਲੀ ਟ੍ਰਾਂਸਜੇਨਿਕਸ ਅਤੇ 179-0 ਸਕੇਲ 'ਤੇ ਅਣ-ਪਰਿਵਰਤਿਤ ਨਿਯੰਤਰਣਾਂ ਵਿੱਚ 4 ਸਵੈ-ਪਰਾਗਣ। ਜ਼ਿਆਦਾਤਰ ਪਰਾਗ ਟਿਊਬਾਂ ਅੰਡਕੋਸ਼ਾਂ ਤੱਕ ਪਹੁੰਚਦੀਆਂ ਹਨ ਸਲੀ ਟ੍ਰਾਂਸਜੇਨਿਕ, ਨਿਯੰਤਰਣ ਦੇ ਸਿਰਫ ਇੱਕ ਬਹੁਤ ਹੀ ਛੋਟੇ ਹਿੱਸੇ ਦੀ ਤੁਲਨਾ ਵਿੱਚ, ਇਹ ਦਰਸਾਉਂਦਾ ਹੈ ਕਿ PGSC0003DMG400016861 ਹੈ ਸਲੀ ਜੀਨ.
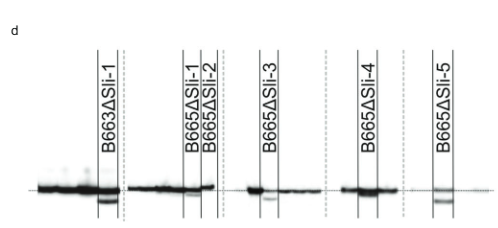
ਅੱਗੇ, ਅਸੀਂ PGSC9DMG0003 (pAGM:CRISPRASli, ਚਿੱਤਰ. 3a). ਅਸੀਂ ਇਸ ਨਿਰਮਾਣ ਨਾਲ ਦੋ SC ਜੀਨੋਟਾਈਪਾਂ (B665 ਅਤੇ B663) ਨੂੰ ਬਦਲਿਆ ਅਤੇ 149 ਪਰਿਵਰਤਿਤ ਪੁਨਰਜਨਮ ਪ੍ਰਾਪਤ ਕੀਤੇ। ਅਸੀਂ ਫਿਰ CRISPR-Cas9-ਪ੍ਰੇਰਿਤ INDELs ਦੀ ਪਛਾਣ ਕਰਨ ਲਈ PAGE ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਨਿਸ਼ਾਨਾ ਐਕਸੋਨ ਦਾ ਵਿਸ਼ਲੇਸ਼ਣ ਕੀਤਾ। ਬਦਕਿਸਮਤੀ ਨਾਲ, pAGM:CRISPRASli ਵੈਕਟਰ ਦੀ ਘੱਟ ਕੁਸ਼ਲਤਾ ਸੀ, 149 ਰੀਜਨਰੇਟਰਾਂ ਵਿੱਚੋਂ ਸਿਰਫ਼ ਛੇ ਨੇ INDEL ਦਿਖਾਏ ਸਲੀ (ਅੰਜੀਰ. 3d). ਇਹਨਾਂ ਵਿੱਚੋਂ ਪੰਜ CRISPR-Cas9 ਲਾਈਨਾਂ INDELs ਲਈ ਵਿਪਰੀਤ ਹਨ, ਪਰ ਇੱਕ ਲਾਈਨ, B665ASli-2, ਇੱਕ ਛੋਟੇ INDEL ਲਈ ਸਮਰੂਪ ਹੈ। ਜਦੋਂ ਕਿ ਬਿਨਾਂ ਪਰਿਵਰਤਿਤ B665 ਆਸਾਨੀ ਨਾਲ ਸਵੈ-ਬੇਰੀਆਂ ਨੂੰ ਸੈੱਟ ਕਰਦਾ ਹੈ ਅਤੇ 105 ਨਿਰੀਖਣ ਕੀਤੀਆਂ ਸ਼ੈਲੀਆਂ ਰਾਹੀਂ ਚੰਗੀ ਸਵੈ-ਪਰਾਗ ਟਿਊਬ ਦਾ ਵਾਧਾ ਦਰਸਾਉਂਦਾ ਹੈ, B665ASli-2 ਸਵੈ-ਪਰਾਗਿਤ ਹੋਣ 'ਤੇ ਬੇਰੀਆਂ ਨੂੰ ਸੈੱਟ ਨਹੀਂ ਕਰਦਾ ਹੈ ਅਤੇ ਇਹ ਪਰਾਗ 78 ਨਿਰੀਖਣ ਕੀਤੀਆਂ ਸ਼ੈਲੀਆਂ (ਚਿੱਤਰ XNUMX) ਦੁਆਰਾ ਵਧਣ ਦੇ ਯੋਗ ਨਹੀਂ ਹੁੰਦਾ ਹੈ। 3b, c ਅਤੇ ਸਪਲੀਮੈਂਟਰੀ ਡੇਟਾ 3), ਹੋਰ ਸਬੂਤ ਪ੍ਰਦਾਨ ਕਰਦੇ ਹੋਏ ਕਿ PGSC0003DMG400016861 ਅਸਲ ਵਿੱਚ ਸਲੀ ਜੀਨ.
S-RNase-ਅਧਾਰਿਤ ਗੇਮਟੋਫਾਈਟਿਕ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਪ੍ਰਣਾਲੀਆਂ ਵਿੱਚ, ਸਵੈ-ਗਰੱਭਧਾਰਣ ਨੂੰ ਪਿਸਟਲ ਦੁਆਰਾ ਪ੍ਰਗਟਾਏ ਗਏ S-RNases ਦੁਆਰਾ ਰੋਕਿਆ ਜਾਂਦਾ ਹੈ ਜੋ ਪਰਾਗ ਟਿਊਬਾਂ ਵਿੱਚ ਦਾਖਲ ਹੁੰਦੇ ਹਨ ਅਤੇ ਸਵੈ-ਪਰਾਗ ਜਾਂ ਕਿਸੇ ਹੋਰ ਪਰਾਗ 'ਤੇ ਸਾਈਟੋਟੌਕਸਿਕ ਪ੍ਰਭਾਵ ਪਾਉਂਦੇ ਹਨ ਜਿਸ ਵਿੱਚ ਮੇਲ ਖਾਂਦੇ S-ਲੋਕਸ ਐੱਫ-ਬਾਕਸ (SLF) ਦੀ ਘਾਟ ਹੁੰਦੀ ਹੈ। ) ਪ੍ਰੋਟੀਨ. ਕ੍ਰਾਸ-ਫਰਟੀਲਾਈਜ਼ੇਸ਼ਨ ਪਰਾਗ-ਪ੍ਰਗਟਾਏ ਗਏ SLF ਪ੍ਰੋਟੀਨ ਦੁਆਰਾ ਸਮਰਥਿਤ ਹੈ ਜੋ ਗੈਰ-ਸਵੈ-ਸ-ਆਰ-ਨੇਜ਼ ਨੂੰ ਪਛਾਣ ਅਤੇ ਡੀਟੌਕਸਫਾਈ ਕਰ ਸਕਦੇ ਹਨ। ਹਰੇਕ S-ਐਲੀਲ ਕਈ SLFs ਨੂੰ ਏਨਕੋਡ ਕਰਦਾ ਹੈ ਜਿਨ੍ਹਾਂ ਵਿੱਚੋਂ ਹਰ ਇੱਕ ਵੱਖਰੇ S-RNase ਨੂੰ ਪਛਾਣ ਸਕਦਾ ਹੈ, ਅਤੇ ਇਕੱਠੇ ਆਲੂ ਵਿੱਚ ਮੌਜੂਦ ਜ਼ਿਆਦਾਤਰ S-RNase ਨੂੰ ਪਛਾਣ ਸਕਦਾ ਹੈ, ਸਿਵਾਏ S-RNase ਨੂੰ ਇੱਕੋ S-ਐਲੀਲ 'ਤੇ ਏਨਕੋਡ ਕੀਤਾ ਗਿਆ ਹੈ। Sli ਇੱਕ F-ਬਾਕਸ ਪ੍ਰੋਟੀਨ PP2-B10 ਨੂੰ ਏਨਕੋਡ ਕਰਦਾ ਹੈ, ਜਿਸ ਵਿੱਚ ਇੱਕ F-ਬਾਕਸ ਡੋਮੇਨ ਹੁੰਦਾ ਹੈ ਜੋ ਇੱਕ ਲੈਕਟਿਨ ਡੋਮੇਨ ਨਾਲ ਜੁੜਿਆ ਹੁੰਦਾ ਹੈ। ਲੈਕਟਿਨ ਡੋਮੇਨ ਕਾਰਬੋਹਾਈਡਰੇਟ ਨਾਲ ਪਰਸਪਰ ਪ੍ਰਭਾਵ ਪਾਉਣ ਲਈ ਜਾਣੇ ਜਾਂਦੇ ਹਨ, ਅਤੇ ਗਲਾਈਕੋਸਾਈਲੇਟਿਡ ਪ੍ਰੋਟੀਨ ਨਾਲ ਗੱਲਬਾਤ ਕਰਨ ਦੇ ਯੋਗ ਹੋ ਸਕਦੇ ਹਨ16. ਇਸ ਤੋਂ ਇਲਾਵਾ, S-RNases ਨੂੰ ਗਲਾਈਕੋਸਾਈਲੇਟਡ ਦਿਖਾਇਆ ਗਿਆ ਹੈ17. ਅਸੀਂ ਇਹ ਅੰਦਾਜ਼ਾ ਲਗਾਉਂਦੇ ਹਾਂ ਕਿ Sli ਦੇ SC ਐਲੀਲ ਦੇ ਪ੍ਰਮੋਟਰ ਵਿੱਚ 533 bp ਸੰਮਿਲਨ ਪਰਾਗ ਵਿੱਚ ਪ੍ਰਗਟਾਵੇ ਨੂੰ ਸਮਰੱਥ ਬਣਾਉਂਦਾ ਹੈ, ਜਿੱਥੇ Sli ਸਵੈ-ਪਰਾਗ ਟਿਊਬ ਦੇ ਵਾਧੇ ਦੀ ਗਿਰਫ਼ਤਾਰੀ ਅਤੇ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਨੂੰ ਗੁਆਉਣ ਲਈ ਸਵੈ-S-RNases ਨੂੰ ਬੰਨ੍ਹਣ ਅਤੇ ਡੀਟੌਕਸੀਫਾਈ ਕਰਨ ਦੇ ਯੋਗ ਹੁੰਦਾ ਹੈ। ਲੈਮਬੀਰ ਦੁਆਰਾ ਆਲੂਆਂ ਵਿੱਚ MITE ਗਤੀਵਿਧੀ ਦੀ ਇੱਕ ਵਿਸਤ੍ਰਿਤ ਜਾਂਚ ਵਿੱਚ ਪਾਇਆ ਗਿਆ ਕਿ ਜੈਨਿਕ ਖੇਤਰਾਂ ਦੇ ਨੇੜੇ MITE ਸੰਮਿਲਨ ਦੇ 2% ਜੀਨ ਸਮੀਕਰਨ ਵਿੱਚ ਤਬਦੀਲੀਆਂ ਨਾਲ ਜੁੜੇ ਹੋਏ ਸਨ।
ਇਸ ਤੋਂ ਇਲਾਵਾ, ਜੈਨਿਕ ਖੇਤਰਾਂ ਦੇ ਨੇੜੇ 1935 ਦੇ ਟੈਸਟ ਕੀਤੇ ਗਏ ਹੈਟ ਸੰਮਿਲਨਾਂ ਵਿੱਚੋਂ, 13 ਦੇ ਨਤੀਜੇ ਵਜੋਂ ਸੰਬੰਧਿਤ ਜੀਨ ਨੂੰ ਅਪਗ੍ਰੇਗੂਲ ਕੀਤਾ ਗਿਆ, ਜੋ ਇਹ ਦਰਸਾਉਂਦਾ ਹੈ ਕਿ ਪਰਾਗ-ਵਿਸ਼ੇਸ਼ ਸਮੀਕਰਨ ਬਦਲਿਆ ਗਿਆ ਹੈ। ਸਲੀ ਅਸਲ ਵਿੱਚ ਇਸਦੇ ਪ੍ਰਮੋਟਰ ਵਿੱਚ 533 ਬੀਪੀ ਸੰਮਿਲਨ ਦੇ ਕਾਰਨ ਹੋ ਸਕਦਾ ਹੈ18. ਫਿਰ ਵੀ, ਇਸ ਪਰਿਕਲਪਨਾ ਦੀ ਵੈਧਤਾ ਨੂੰ ਨਿਰਧਾਰਤ ਕਰਨ ਲਈ ਹੋਰ ਖੋਜ ਦੀ ਲੋੜ ਹੈ।
ਪਹਿਲਾਂ, ਕ੍ਰੋਮੋਸੋਮ 333 'ਤੇ 12 kb ਅੰਤਰਾਲ ਦੀ ਪਛਾਣ ਕਰਨ ਲਈ ਕਲੌਟ ਐਟ ਅਲ ਨੇ ਇੱਕ ਬਲਕਡ ਸੇਗਰੀਗੈਂਟ ਕੇ-ਮੇਰ ਮੈਪਿੰਗ ਪਹੁੰਚ ਦੀ ਵਰਤੋਂ ਕੀਤੀ ਜਿਸ ਵਿੱਚ Sli ਸਥਿਤ ਹੋਣਾ ਚਾਹੀਦਾ ਹੈ।19. ਇੱਥੇ, ਅਸੀਂ ਇੱਕ F12 ਆਬਾਦੀ ਵਿੱਚ ਕ੍ਰੋਮੋਸੋਮ 1 ਦੇ ਉਸੇ ਖੇਤਰ ਵਿੱਚ ਸਲੀ ਲੋਕਸ ਨੂੰ ਮੈਪ ਕੀਤਾ ਅਤੇ 12.6 ਜੀਨਾਂ ਵਾਲੇ ਅੰਤਰਾਲ ਨੂੰ 2 KB ਤੱਕ ਘਟਾਉਣ ਲਈ ਇੱਕ ਰੀਕੌਂਬੀਨੈਂਟ ਸਕ੍ਰੀਨਿੰਗ ਦੀ ਵਰਤੋਂ ਕੀਤੀ। ਸਮੀਕਰਨ ਵਿਸ਼ਲੇਸ਼ਣ ਤੋਂ ਪਤਾ ਲੱਗਾ ਹੈ ਕਿ ਇਹਨਾਂ ਜੀਨਾਂ ਵਿੱਚੋਂ ਇੱਕ ਦਾ SC ਐਲੀਲ ਵਿਸ਼ੇਸ਼ ਤੌਰ 'ਤੇ SC ਜੀਨੋਟਾਈਪਾਂ ਤੋਂ ਪਰਾਗ ਵਿੱਚ ਪ੍ਰਗਟ ਹੁੰਦਾ ਹੈ। ਅੰਤ ਵਿੱਚ, ਟ੍ਰਾਂਸਜੇਨਿਕ ਸਮੀਕਰਨ ਅਤੇ CRISPR-Cas9-ਪ੍ਰੇਰਿਤ ਨਾਕ-ਆਊਟ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ, ਅਸੀਂ ਸਿੱਟੇ ਵਜੋਂ ਦਿਖਾਉਂਦੇ ਹਾਂ ਕਿ PGSC0003DMG400016861 Sli ਹੈ। ਜਦੋਂ ਕਿ ਲੇਮਬੀਰ ਦੁਆਰਾ ਕੀਤੇ ਅਧਿਐਨ ਨੇ ਦਿਖਾਇਆ ਹੈ ਕਿ MITEs ਟਿਸ਼ੂ-ਵਿਸ਼ੇਸ਼ ਤਰੀਕੇ ਨਾਲ ਨਜ਼ਦੀਕੀ ਜੀਨਾਂ ਨੂੰ ਉੱਚਿਤ ਕਰ ਸਕਦੇ ਹਨ, ਇਹ ਸਾਬਤ ਕਰਨ ਲਈ ਹੋਰ ਖੋਜ ਦੀ ਲੋੜ ਹੈ ਕਿ Sli ਪ੍ਰਮੋਟਰ ਵਿੱਚ MITE ਦੀ ਮੌਜੂਦਗੀ ਇਸਦੇ ਪਰਾਗ-ਵਿਸ਼ੇਸ਼ ਸਮੀਕਰਨ ਦਾ ਕਾਰਨ ਹੈ।
ਇਸ ਅਧਿਐਨ (ਪੂਰਕ ਡੇਟਾ 4) ਵਿੱਚ ਜਾਂਚ ਕੀਤੀ ਸਮੱਗਰੀ ਵਿੱਚ, ਅਸੀਂ Sli ਲਈ ਸਮਰੂਪਤਾ ਨਾਲ ਸੰਬੰਧਿਤ ਘਾਤਕਤਾ ਦੀਆਂ ਪਿਛਲੀਆਂ ਰਿਪੋਰਟਾਂ ਦੀ ਪੁਸ਼ਟੀ ਨਹੀਂ ਕਰ ਸਕੇ, ਕਿਉਂਕਿ ਸਾਨੂੰ Sli ਲਈ ਵਿਹਾਰਕ F2 ਪੌਦਿਆਂ ਨੂੰ ਸਮਰੂਪਤਾ ਦਾ ਪਤਾ ਲੱਗਾ ਹੈ ਜੋ ਬੇਰੀਆਂ ਨੂੰ ਸੈੱਟ ਕਰਨ ਦੇ ਯੋਗ ਸਨ (ਪੂਰਕ ਡੇਟਾ 1)12>20. ਇਸ ਤੋਂ ਇਲਾਵਾ, ਇਨਬ੍ਰੇਡ-ਲਾਈਨ ਸੋਲਿਨਟਸ ਦੇ ਜੀਨੋਮ ਕ੍ਰਮ, ਅਤੇ ਨਾਲ ਹੀ ਇਨਬ੍ਰੇਡ-ਲਾਈਨ M6, ਇਹ ਦਰਸਾਉਂਦੇ ਹਨ ਕਿ ਇਹ ਦੋਵੇਂ ਲਾਈਨਾਂ ਸਲੀ ਲਈ ਸਮਰੂਪ ਹਨ, ਇਹ ਦਰਸਾਉਂਦੀਆਂ ਹਨ ਕਿ ਸਲੀ ਲਈ ਸਮਰੂਪਤਾ ਘਾਤਕ ਨਹੀਂ ਹੈ, ਹਾਲਾਂਕਿ ਇਹ ਅਜੇ ਵੀ ਸੰਭਵ ਹੈ ਕਿ ਇੱਕ ਘਾਤਕ ਪੂਰਵਜ ਵਿੱਚ Sli ਨਾਲ ਜੈਨੇਟਿਕ ਤੌਰ 'ਤੇ ਜੁੜੇ ਐਲੀਲ ਨੂੰ ਇਹਨਾਂ ਜੀਨੋਟਾਈਪਾਂ ਵਿੱਚ ਪੁਨਰ-ਸੰਯੋਜਨ ਦੁਆਰਾ ਹਟਾ ਦਿੱਤਾ ਗਿਆ ਹੈ21'22. ਹਾਲਾਂਕਿ, ਇਸ ਅਧਿਐਨ ਵਿੱਚ ਤਿਆਰ ਕੀਤੇ ਗਏ ਡੇਟਾ ਤੋਂ, ਅਸੀਂ ਇਸ ਸੰਭਾਵਨਾ ਨੂੰ ਬਾਹਰ ਨਹੀਂ ਕੱਢ ਸਕਦੇ ਕਿ F2 ਆਬਾਦੀ ਵਿੱਚ ਦੇਖਿਆ ਗਿਆ ਵੱਖਰਾ ਵਿਗਾੜ Sli ਦੇ SI ਐਲੀਲ ਨਾਲ ਪੜਾਅ ਵਿੱਚ ਜੁੜੇ ਇੱਕ ਘਾਤਕ ਐਲੀਲ ਕਾਰਨ ਹੋਇਆ ਹੈ। ਅਜੇ ਤੱਕ, ਇਹ ਅਸਪਸ਼ਟ ਹੈ ਕਿ ਕੀ Sli ਖੁਦ S-RNases ਨੂੰ ਸਿੱਧੇ ਤੌਰ 'ਤੇ ਪਛਾਣ ਅਤੇ ਡੀਟੌਕਸੀਫਾਈ ਕਰ ਸਕਦਾ ਹੈ। ਇਸ ਤੋਂ ਇਲਾਵਾ, ਇਹ ਸਪੱਸ਼ਟ ਨਹੀਂ ਹੈ ਕਿ ਕੀ Sli ਦੇ ਨਤੀਜੇ ਵਜੋਂ ਸਾਰੇ S-ਲੋਕਸ ਜੀਨੋਟਾਈਪਾਂ ਵਿੱਚ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਹੁੰਦੀ ਹੈ। ਇਹ ਸੰਭਵ ਹੈ ਕਿ ਕੁਝ ਐਸ-ਐਲੀਲਾਂ ਦੇ ਕੰਮ ਨੂੰ Sli ਦੁਆਰਾ ਰੋਕਿਆ ਨਹੀਂ ਜਾ ਸਕਦਾ ਹੈ। ਇਸ ਸਵਾਲ ਨੂੰ ਹੱਲ ਕਰਨ ਲਈ ਹੋਰ ਖੋਜ ਦੀ ਲੋੜ ਹੈ।
ਜਦੋਂ ਕਿ Sli ਦੀ ਪਛਾਣ ਡਿਪਲੋਇਡ ਆਲੂ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਇਨਬ੍ਰੇਡ-ਲਾਈਨ-ਅਧਾਰਤ ਹਾਈਬ੍ਰਿਡ ਪ੍ਰਜਨਨ ਨੂੰ ਸਮਰੱਥ ਬਣਾਉਂਦੀ ਹੈ, ਹੋਰ ਰੁਕਾਵਟਾਂ ਬਾਕੀ ਹਨ। ਸਭ ਤੋਂ ਪਹਿਲਾਂ, ਸਭ ਤੋਂ ਮਹੱਤਵਪੂਰਨ ਤੌਰ 'ਤੇ, ਡਿਪਲੋਇਡ ਆਲੂ ਇਨਬ੍ਰੀਡਿੰਗ ਡਿਪਰੈਸ਼ਨ ਤੋਂ ਪੀੜਤ ਹੈ, ਜਿਸ ਨਾਲ ਪ੍ਰਜਨਨ ਦੇ ਬਾਅਦ ਜੋਸ਼ ਅਤੇ ਉਪਜਾਊ ਸ਼ਕਤੀ ਘੱਟ ਜਾਂਦੀ ਹੈ। ਡਿਪਲੋਇਡ ਆਲੂ ਲਾਈਨਾਂ ਦੇ ਨਿਰੰਤਰ ਸਵੈ-ਫਰਟੀਲਾਈਜ਼ੇਸ਼ਨ ਦੁਆਰਾ ਨੁਕਸਾਨਦੇਹ ਐਲੀਲਾਂ ਨੂੰ ਸਾਫ਼ ਕਰਨਾ ਪ੍ਰਜਨਨ ਉਦਾਸੀ ਨੂੰ ਘਟਾਉਣ ਦਾ ਇੱਕ ਕੁਸ਼ਲ ਤਰੀਕਾ ਹੈ ਅਤੇ ਇਸਦੇ ਨਤੀਜੇ ਵਜੋਂ ਪਹਿਲਾਂ ਹੀ ਤੁਲਨਾਤਮਕ ਤੌਰ 'ਤੇ ਜੋਸ਼ਦਾਰ ਅਤੇ ਉਪਜਾਊ ਆਲੂਆਂ ਦੀਆਂ ਰੇਖਾਵਾਂ ਪੈਦਾ ਹੋ ਗਈਆਂ ਹਨ।2'23-25. ਦੂਜਾ, ਇਸ ਅਧਿਐਨ ਵਿੱਚ ਵਰਤੀ ਗਈ Sli ਦਾ ਦਾਨੀ, DS, ਇੱਕ ਐਸ ਤੋਂ ਲਿਆ ਗਿਆ ਹੈ. chacoense ਐਕਸੈਸ਼ਨ, ਸੰਭਾਵੀ ਤੌਰ 'ਤੇ ਐਸ ਤੋਂ ਨੁਕਸਾਨਦੇਹ ਐਲੀਲਾਂ ਦੇ ਲਿੰਕੇਜ ਡਰੈਗ ਨਾਲ ਸਮੱਸਿਆਵਾਂ ਵੱਲ ਅਗਵਾਈ ਕਰਦਾ ਹੈ. chacoense. ਸੋਲਿੰਟਾ ਦੇ ਡਿਪਲੋਇਡ ਬ੍ਰੀਡਿੰਗ ਪ੍ਰੋਗਰਾਮ ਵਿੱਚ, ਸਾਨੂੰ ਐਸ. ਚੈਕੋਏਂਸ ਤੋਂ ਆਉਣ ਵਾਲੇ ਨੁਕਸਾਨਦੇਹ ਐਲੀਲਾਂ ਦੇ ਕਾਰਨ ਸਪੱਸ਼ਟ ਸਮੱਸਿਆਵਾਂ ਨਹੀਂ ਦਿਖਾਈ ਦਿੰਦੀਆਂ। ਇਸ ਤੋਂ ਇਲਾਵਾ, ਸਵੈ-ਅਨੁਕੂਲਤਾ 'ਤੇ ਇੱਕ ਤਾਜ਼ਾ ਅਧਿਐਨ ਨੇ ਇਹ ਖੁਲਾਸਾ ਕੀਤਾ ਹੈ ਕਿ SC-ਵਿਸ਼ੇਸ਼ k-mers ਪਹਿਲਾਂ ਤੋਂ ਹੀ ਕਈ ਟੈਟਰਾਪਲੋਇਡ ਕਿਸਮਾਂ ਵਿੱਚ ਮੌਜੂਦ ਹਨ, ਜੋ ਕਿ ਇਹਨਾਂ ਕਿਸਮਾਂ ਤੋਂ ਤਿਆਰ ਕੀਤੇ ਗਏ ਡਾਈਹਾਪਲੋਇਡਾਂ ਨੂੰ ਸਲਾਈ ਡੋਨਰ ਵਜੋਂ ਵਰਤ ਕੇ ਇਸ ਸੰਭਾਵੀ ਲਿੰਕੇਜ ਡਰੈਗ ਨੂੰ ਪੂਰੀ ਤਰ੍ਹਾਂ ਰੋਕਣ ਲਈ ਇੱਕ ਰਸਤਾ ਪ੍ਰਦਾਨ ਕਰਦੇ ਹਨ।19.
ਢੰਗ
ਪੌਦਾ ਸਮੱਗਰੀ. ਸਾਰੀਆਂ ਵਰਤੀਆਂ ਜਾਣ ਵਾਲੀਆਂ ਪੌਦਿਆਂ ਦੀਆਂ ਸਮੱਗਰੀਆਂ ਨੂੰ ਸਪਲੀਮੈਂਟਰੀ ਡੇਟਾ 4 ਵਿੱਚ ਸੂਚੀਬੱਧ ਕੀਤਾ ਗਿਆ ਹੈ
ਗ੍ਰੀਨਹਾਉਸ ਹਾਲਾਤ. ਸਾਰੇ ਪੌਦੇ ਗ੍ਰੀਨਹਾਉਸਾਂ ਵਿੱਚ ਉਗਾਏ ਗਏ ਸਨ ਜੋ ਤਾਪਮਾਨ 14 ਡਿਗਰੀ ਸੈਲਸੀਅਸ ਤੋਂ ਹੇਠਾਂ ਡਿੱਗਣ 'ਤੇ ਗਰਮ ਕੀਤੇ ਜਾਂਦੇ ਸਨ ਅਤੇ ਜਦੋਂ ਤਾਪਮਾਨ 19 ਡਿਗਰੀ ਸੈਲਸੀਅਸ ਤੋਂ ਉੱਪਰ ਵੱਧ ਜਾਂਦਾ ਸੀ ਤਾਂ ਖਿੜਕੀਆਂ ਖੋਲ੍ਹ ਕੇ ਠੰਡਾ ਕੀਤਾ ਜਾਂਦਾ ਸੀ। ਜਦੋਂ ਰੋਸ਼ਨੀ ਦੀ ਤੀਬਰਤਾ 85 W/M2 ਤੋਂ ਘੱਟ ਜਾਂਦੀ ਹੈ ਤਾਂ ਨਕਲੀ ਰੋਸ਼ਨੀ ਕੁਦਰਤੀ ਰੋਸ਼ਨੀ ਨੂੰ ਪੂਰਕ ਕਰਦੀ ਹੈ। ਪੌਦਿਆਂ ਨੂੰ ਲੈਂਸੇ ਪੋਟਗ੍ਰਾਂਡ (ਲੈਂਟਸੇ ਪੋਟਗ੍ਰਾਂਡ ਬੀਵੀ, ਕੈਟਵਿਜਕ, ਨੀਦਰਲੈਂਡਜ਼) ਤੋਂ ਇੱਕ ਵਿਸ਼ੇਸ਼ ਆਲੂ ਸਬਸਟਰੇਟ ਮਿਸ਼ਰਣ ਵਿੱਚ ਉਗਾਇਆ ਗਿਆ ਸੀ। ਲੋੜੀਂਦੇ pH ਪੱਧਰ ਨੂੰ ਯਕੀਨੀ ਬਣਾਉਣ ਲਈ ਵਰਤਿਆ ਜਾਣ ਵਾਲਾ ਸਬਸਟਰੇਟ ਮਿਸ਼ਰਣ ਸੰਤੁਲਿਤ ਪਾਣੀ ਦੇ ਗ੍ਰਹਿਣ, ਬੁਨਿਆਦੀ ਹੌਲੀ-ਰਿਲੀਜ਼ ਖਾਦ, ਅਤੇ ਚੂਨੇ ਲਈ ਇੱਕ ਪੀਟ ਮਿਸ਼ਰਣ ਤੋਂ ਬਣਿਆ ਹੈ। ਸਬਸਟਰੇਟ ਮਿਸ਼ਰਣ ਨੂੰ 20:20:20 ਨਾਈਟ੍ਰੋਜਨ: ਫਾਸਫੋਰਸ: ਪੋਟਾਸ਼ੀਅਮ ਘੋਲ 1.5 ਦੀ ਬਿਜਲਈ ਚਾਲਕਤਾ (EC) ਨਾਲ ਖਾਦ ਬਣਾਇਆ ਗਿਆ ਸੀ।
ਸਵੈ-ਅਨੁਕੂਲਤਾ ਦਾ ਮੁਲਾਂਕਣ. ਫੁੱਲਾਂ ਅਤੇ ਮੁਕੁਲਾਂ ਨੂੰ ਹਫ਼ਤੇ ਵਿੱਚ ਇੱਕ ਵਾਰ ਗਿਣਿਆ ਜਾਂਦਾ ਸੀ ਅਤੇ ਜੋਸ਼ ਹਰ ਮਹੀਨੇ ਇੱਕ ਵਾਰ 1 ਤੋਂ 9 ਦੇ ਪੈਮਾਨੇ 'ਤੇ ਅੰਕਿਤ ਕੀਤਾ ਜਾਂਦਾ ਸੀ, 1 ਇੱਕ ਬਹੁਤ ਹੀ ਗੈਰ-ਜੋਸ਼ਦਾਰ ਪੌਦਾ ਸੀ, ਅਤੇ 9 ਇੱਕ ਬਹੁਤ ਹੀ ਜੋਸ਼ਦਾਰ ਪੌਦਾ ਸੀ। ਇੱਕ ਪੌਦੇ ਦੇ ਇੱਕ ਤੋਂ ਵੱਧ ਫੁੱਲਾਂ ਦੇ ਪਰਾਗ ਨੂੰ ਇੱਕ ਏਪੇਨਡੋਰਫ ਟਿਊਬ ਵਿੱਚ ਇਕੱਠਾ ਕੀਤਾ ਗਿਆ ਸੀ ਅਤੇ ਉਸੇ ਫੁੱਲਾਂ 'ਤੇ ਸਵੈ-ਪਰਾਗੀਕਰਨ ਲਈ ਤੁਰੰਤ ਵਰਤਿਆ ਜਾਂਦਾ ਸੀ ਜਿਸ ਵਿੱਚ ਪ੍ਰਤੀ ਪੌਦਾ ਪ੍ਰਤੀ ਹਫ਼ਤੇ ਵੱਧ ਤੋਂ ਵੱਧ 10 ਫੁੱਲ ਹੁੰਦੇ ਹਨ। ਉਹ ਪੌਦੇ ਜੋ ਪ੍ਰਤੀ ਸਵੈ-ਬੇਰੀ ਘੱਟੋ-ਘੱਟ 35 ਬੀਜਾਂ ਵਾਲੇ ਦੋ ਤੋਂ ਵੱਧ ਸਵੈ-ਬੇਰੀ ਸੈੱਟ ਕਰਦੇ ਹਨ, ਸਵੈ-ਅਨੁਕੂਲ ਵਜੋਂ ਸ਼੍ਰੇਣੀਬੱਧ ਕੀਤੇ ਗਏ ਸਨ। ਮਾਦਾ ਉਪਜਾਊ ਸ਼ਕਤੀ ਨੂੰ ਨਿਰਧਾਰਤ ਕਰਨ ਲਈ, ਪੌਦਿਆਂ ਨੂੰ ਘੱਟੋ-ਘੱਟ ਤਿੰਨ ਗੈਰ-ਸੰਬੰਧਿਤ ਜੀਨੋਟਾਈਪਾਂ ਤੋਂ ਵੱਡੇ ਪਰਾਗ ਨਾਲ ਪਰਾਗਿਤ ਕੀਤਾ ਗਿਆ ਸੀ। ਉਹ ਪੌਦੇ ਜਿਨ੍ਹਾਂ ਨੇ ਘੱਟੋ-ਘੱਟ 10 ਸਵੈ-ਪਰਾਗਿਤ ਕਰਨ ਤੋਂ ਬਾਅਦ ਸਵੈ-ਬੇਰੀਆਂ ਨੂੰ ਸੈੱਟ ਨਹੀਂ ਕੀਤਾ, ਪਰ ਘੱਟੋ-ਘੱਟ ਇੱਕ ਬਲਕ ਬੇਰੀ ਸੈੱਟ ਕੀਤੀ ਅਤੇ ਸਵੈ-ਪਰਾਗਿਤ ਸ਼ੈਲੀਆਂ ਦੇ ਸੂਖਮ ਵਿਸ਼ਲੇਸ਼ਣ ਵਿੱਚ ਉਪਜਾਊ ਪਰਾਗ ਨੂੰ ਦਿਖਾਇਆ, ਉਹਨਾਂ ਨੂੰ ਸਵੈ-ਅਸੰਗਤ ਵਜੋਂ ਸ਼੍ਰੇਣੀਬੱਧ ਕੀਤਾ ਗਿਆ। ਮੈਪਿੰਗ ਆਬਾਦੀ ਤੋਂ 40 ਜੀਨੋਟਾਈਪਾਂ ਲਈ ਜਿਸ ਵਿੱਚ ਬੇਰੀ ਅਤੇ ਬੀਜ ਸੈੱਟ ਡੇਟਾ ਨਿਰਣਾਇਕ ਸਨ (17SC11: n = 14, 18SC11: n = 7, 18SC12: n = 19), phenotypical ਵਰਗੀਕਰਣ ਸਟਾਈਲ ਦੁਆਰਾ ਸਵੈ-ਪਰਾਗ ਟਿਊਬ ਵਿਕਾਸ 'ਤੇ ਅਧਾਰਤ ਸੀ (ਸਪਲੀਮੈਂਟਰੀ ਡੇਟਾ 1 ਵਿੱਚ ਨੋਟ ਕੀਤਾ ਗਿਆ ਹੈ)।
ਸ਼ੈਲੀ ਇਮੇਜਿੰਗ. ਪਰਾਗ ਟਿਊਬ ਦੇ ਵਾਧੇ ਦੀ ਕਲਪਨਾ ਕਰਨ ਲਈ, ਪਰਾਗਿਤ ਸਟਾਈਲ ਨੂੰ ਪਰਾਗਿਤ ਕਰਨ ਤੋਂ 24-48 ਘੰਟੇ ਬਾਅਦ ਹਟਾ ਦਿੱਤਾ ਗਿਆ ਅਤੇ ਫਿਰ 3:1 ਈਥਾਨੋਲ: ਐਸੀਟਿਕ ਐਸਿਡ ਵਿੱਚ ਘੱਟੋ-ਘੱਟ 24 ਘੰਟਿਆਂ ਲਈ ਫਿਕਸ ਕੀਤਾ ਗਿਆ। ਫਿਰ ਸਟਾਈਲ ਨੂੰ 8M NaOH ਵਿੱਚ 10 °C 'ਤੇ 65 ਮਿੰਟਾਂ ਲਈ ਮੈਸਰੇਟ ਕੀਤਾ ਗਿਆ ਅਤੇ ਡੀਓਨਾਈਜ਼ਡ ਪਾਣੀ ਨਾਲ ਦੋ ਵਾਰ ਕੁਰਲੀ ਕੀਤਾ ਗਿਆ। ਸਟਾਈਲਾਂ ਨੂੰ ਮਾਈਕ੍ਰੋਸਕੋਪੀ ਸਲਾਈਡਾਂ 'ਤੇ ਰੱਖਿਆ ਗਿਆ ਸੀ ਅਤੇ 2MK ਵਿੱਚ 5% ਐਨੀਲਾਈਨ ਬਲੂ (ਕਾਰਲ ਰੋਥ GmbH) ਦੀ ਵਰਤੋਂ ਕਰਕੇ 0.1-0.1 ਮਿੰਟ ਲਈ ਦਾਗਿਆ ਗਿਆ ਸੀ।4P2O7 (pH = 7), ਫਿਰ ਇੱਕ ਕਵਰਸਲਿਪ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਗਲਾਈਸਰੋਲ ਵਿੱਚ ਕੁਚਲਿਆ ਗਿਆ ਅਤੇ ਇੱਕ ਫਿਲਟਰ ਸੈੱਟ 01 (BP 365/12, FT 395 ਅਤੇ LP 397) ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਇੱਕ Zeiss Axiolab ਫਲੋਰੋਸੈਂਸ ਮਾਈਕ੍ਰੋਸਕੋਪ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਦੇਖਿਆ ਗਿਆ। ਸਾਰੀਆਂ ਸ਼ੈਲੀਆਂ ਨੂੰ ਦੋ ਮਾਪਦੰਡਾਂ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਦੇਖਿਆ ਅਤੇ ਸਕੋਰ ਕੀਤਾ ਗਿਆ ਸੀ: (1) ਸ਼ੈਲੀ ਵਿੱਚ ਸਭ ਤੋਂ ਡੂੰਘੀ ਪ੍ਰਵੇਸ਼, ਜਿਵੇਂ ਕਿ ਅਧਿਕਤਮ ਪ੍ਰਵੇਸ਼ ਦੀ ਪ੍ਰਤੀਸ਼ਤਤਾ ਵਿੱਚ ਦਰਸਾਇਆ ਗਿਆ ਹੈ, (2) ਪਰਾਗ ਟਿਊਬਾਂ ਦਾ % ਡੂੰਘੀ ਪ੍ਰਵੇਸ਼ ਤੱਕ ਪਹੁੰਚਣਾ। ਫਿਰ ਅਸੀਂ ਇਹਨਾਂ ਪ੍ਰਤੀਸ਼ਤਾਂ ਨੂੰ 0-4 ਪੈਮਾਨੇ ਵਿੱਚ ਬਦਲ ਦਿੱਤਾ, ਜਿੱਥੇ ਉਹ ਸਟਾਈਲ ਜਿਹਨਾਂ ਵਿੱਚ ਕੋਈ ਪਰਾਗ ਟਿਊਬਾਂ ਅੰਡਾਸ਼ਯ ਤੱਕ ਨਹੀਂ ਪਹੁੰਚਦੀਆਂ ਸਨ, ਉਹਨਾਂ ਨੂੰ 0 ਦਾ ਸਕੋਰ ਮਿਲਦਾ ਸੀ, ਉਹਨਾਂ ਸਟਾਈਲ ਜਿਹਨਾਂ ਵਿੱਚ ਪਰਾਗ ਟਿਊਬਾਂ ਦੇ 0 ਤੋਂ 25% ਵਿਚਕਾਰ 1 ਦਾ ਸਕੋਰ ਪ੍ਰਾਪਤ ਹੁੰਦਾ ਸੀ, ਉਹਨਾਂ ਸਟਾਈਲ ਜਿਹਨਾਂ ਵਿੱਚ 25 ਦੇ ਵਿਚਕਾਰ ਅਤੇ ਅੰਡਾਸ਼ਯ ਤੱਕ ਪਹੁੰਚੀਆਂ 50% ਪਰਾਗ ਟਿਊਬਾਂ ਨੂੰ 2 ਦਾ ਸਕੋਰ ਮਿਲਿਆ, ਸਟਾਈਲ ਜਿਸ ਵਿੱਚ 50 ਤੋਂ 75% ਦੇ ਵਿਚਕਾਰ ਪਰਾਗ ਟਿਊਬਾਂ ਅੰਡਾਸ਼ਯ ਵਿੱਚ ਪਹੁੰਚੀਆਂ, ਉਹਨਾਂ ਨੂੰ 3 ਦਾ ਸਕੋਰ ਮਿਲਿਆ, ਅਤੇ ਸਟਾਈਲ ਜਿਹਨਾਂ ਵਿੱਚ 75% ਤੋਂ ਵੱਧ ਪਰਾਗ ਟਿਊਬਾਂ ਅੰਡਾਸ਼ਯ ਵਿੱਚ ਪਹੁੰਚੀਆਂ। 4 ਦਾ ਸਕੋਰ।
ਚਿੱਤਰ ਪ੍ਰਾਪਤੀ. ਚੁਣੀਆਂ ਗਈਆਂ ਸ਼ੈਲੀਆਂ ਨੂੰ ਫਿਲਟਰ ਸੈੱਟ 01 ਦੇ ਨਾਲ ਇੱਕ Zeiss AxioCam ICc 5 ਦੇ ਨਾਲ ਇੱਕ Zeiss Axiophot ਮਾਈਕ੍ਰੋਸਕੋਪ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਚਿੱਤਰਿਆ ਗਿਆ ਸੀ। ਚਿੱਤਰ Zeiss Zen 2.3 (ਨੀਲਾ ਐਡੀਸ਼ਨ) ਸਾਫਟਵੇਅਰ ਪੈਕੇਜ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਬਣਾਏ ਗਏ ਸਨ। ਪ੍ਰਾਪਤੀ ਦੇ ਦੌਰਾਨ, ਬੈਕਗ੍ਰਾਉਂਡ ਨੂੰ ਘੱਟ ਤੋਂ ਘੱਟ ਕਰਨ ਲਈ ਸੈਟਿੰਗਾਂ ਨੂੰ ਐਡਜਸਟ ਕੀਤਾ ਗਿਆ ਸੀ। ਸਟਾਈਲਾਂ ਨੂੰ x5 ਉਦੇਸ਼ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਚਿੱਤਰ ਬਣਾਇਆ ਗਿਆ ਸੀ ਅਤੇ 2464-ਬਿੱਟ ਡੂੰਘਾਈ ਦੇ ਨਾਲ 2056 x 24 ਪਿਕਸਲ ਦੇ ਰੈਜ਼ੋਲਿਊਸ਼ਨ ਨਾਲ TIFF ਫਾਈਲਾਂ ਵਜੋਂ ਸੁਰੱਖਿਅਤ ਕੀਤਾ ਗਿਆ ਸੀ। ਫਿਰ ਪਨਵਿਊ ਚਿੱਤਰ ਅਸੈਂਬਲਰ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਅੱਠ ਵੱਖ-ਵੱਖ ਚਿੱਤਰਾਂ ਨੂੰ ਇਕੱਠਾ ਕੀਤਾ ਗਿਆ ਸੀ। ਅੰਜੀਰ ਬਣਾਉਣ ਲਈ ਇਕੱਠੇ ਕੀਤੇ ਸਟਾਈਲ ਦੇ ਉਲਟ ਅਤੇ ਚਮਕ ਨੂੰ ਐਡਜਸਟ ਕੀਤਾ ਗਿਆ ਸੀ। 1a ਅਤੇ 3c.
ਡੀਐਨਏ ਕੱਢਣ. ਮੈਪਿੰਗ ਆਬਾਦੀ ਦੇ KASP ਵਿਸ਼ਲੇਸ਼ਣ ਲਈ, ਨਿਰਮਾਤਾ ਦੁਆਰਾ ਸਪਲਾਈ ਕੀਤੇ ਗਏ ਪ੍ਰੋਟੋਕੋਲ ਦੇ ਅਨੁਸਾਰ sbeadex™ ਕਿੱਟਾਂ (LGC Genomics GmbH, ਬਰਲਿਨ, ਜਰਮਨੀ) ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ DNA ਕੱਢਣ ਲਈ ਪੱਤਿਆਂ ਦੇ ਨਮੂਨੇ VHLGenetics (Wageningen, The Netherlands) ਨੂੰ ਭੇਜੇ ਗਏ ਸਨ।
KASP ਵਿਸ਼ਲੇਸ਼ਣ. ਮੁਕਾਬਲਾਤਮਕ ਐਲੀਲ-ਵਿਸ਼ੇਸ਼ PCR (KASP™) ਵਿਸ਼ਲੇਸ਼ਣ VHLGenetics (Wageningen, The Netherlands) ਦੁਆਰਾ KASP ਅਸੇਸ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਕੀਤਾ ਗਿਆ ਸੀ ਜੋ ਸਾਡੀ ਸਮੱਗਰੀ ਵਿੱਚ ਵੱਖ ਹੋਣ ਵਾਲੇ SNPs ਲਈ ਖਾਸ ਹੋਣ ਲਈ ਤਿਆਰ ਕੀਤੇ ਗਏ ਸਨ। KASP ਅਸੈਸ ਨਿਰਮਾਤਾ (LGC Genomics GmbH, ਬਰਲਿਨ, ਜਰਮਨੀ) ਦੁਆਰਾ ਪ੍ਰਦਾਨ ਕੀਤੇ ਗਏ ਪ੍ਰੋਟੋਕੋਲ ਦੇ ਅਨੁਸਾਰ ਕੀਤੇ ਗਏ ਸਨ। KASP ਅਸੈਸ ਦੇ ਨਤੀਜਿਆਂ ਨੂੰ SNPviewer (lgcgroup.com/products/genotyping-software/snpviewer 'ਤੇ ਉਪਲਬਧ) ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਸਹੀ ਅਲੱਗ-ਥਲੱਗ ਅਤੇ ਜੀਨੋਟਾਈਪ ਕਾਲਿੰਗ ਦੀ ਪੁਸ਼ਟੀ ਕਰਨ ਲਈ ਵਿਜ਼ੂਅਲ ਕੀਤਾ ਗਿਆ ਸੀ।
ਲਿੰਕੇਜ ਵਿਸ਼ਲੇਸ਼ਣ. ਸਵੈ-ਅਨੁਕੂਲ ਮਾਦਾ ਮਾਪਿਆਂ ਦੀਆਂ ਹੈਪਲੋਟਾਈਪਾਂ ਨੂੰ ਜੀਨੋਟਾਈਪ ਡੇਟਾ ਤੋਂ ਵੱਖ-ਵੱਖ SNPs ਵਿਚਕਾਰ ਪੁਨਰ-ਸੰਯੋਜਨ ਦਰਾਂ ਦਾ ਵਿਸ਼ਲੇਸ਼ਣ ਕਰਕੇ ਪੁਨਰਗਠਨ ਕੀਤਾ ਗਿਆ ਸੀ। ਇਹ ਡੇਟਾ SNP ਕਾਲਾਂ ਨੂੰ ਇੱਕ "axb" ਫਾਰਮੈਟ ਵਿੱਚ ਬਦਲਣ ਲਈ ਵਰਤਿਆ ਗਿਆ ਸੀ, ਜਿਸ ਵਿੱਚ "a" ਹੈਪਲੋਟਾਈਪ Sli ਦੇ ਸਵੈ-ਅਨੁਕੂਲ ਐਲੀਲ ਨਾਲ ਜੁੜਿਆ ਹੋਇਆ ਹੈ, ਜਦੋਂ ਕਿ "b" ਹੈਪਲੋਟਾਈਪ Sli ਦੇ ਇੱਕ ਸਵੈ-ਅਨੁਕੂਲ ਐਲੀਲ ਨਾਲ ਜੁੜਿਆ ਹੋਇਆ ਹੈ। ਲਿੰਕੇਜ ਨਕਸ਼ੇ Joinmap 4.1 ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਬਣਾਏ ਗਏ ਸਨ26 ਆਬਾਦੀ ਦੀ ਕਿਸਮ DH ਅਤੇ ਡਿਫੌਲਟ ਸੈਟਿੰਗਾਂ ਦੇ ਨਾਲ।
QTL ਮੈਪਿੰਗ. ਫੀਨੋਟਾਈਪ ਡੇਟਾ ਨੂੰ ਹਰੇਕ ਸਵੈ-ਅਨੁਕੂਲ ਜੀਨੋਟਾਈਪ ਨੂੰ 1, ਹਰੇਕ ਸਵੈ-ਅਨੁਕੂਲ ਜੀਨੋਟਾਈਪ ਨੂੰ 0, ਅਤੇ * ਉਹਨਾਂ ਜੀਨੋਟਾਈਪਾਂ ਲਈ ਨਿਰਧਾਰਤ ਕਰਕੇ ਇੱਕ ਸੰਖਿਆਤਮਕ ਵਿਸ਼ੇਸ਼ਤਾ ਵਿੱਚ ਬਦਲਿਆ ਗਿਆ ਸੀ ਜਿਨ੍ਹਾਂ ਲਈ ਅਨੁਕੂਲਤਾ ਨਿਰਧਾਰਤ ਨਹੀਂ ਕੀਤੀ ਜਾ ਸਕਦੀ ਸੀ। QTL ਮੈਪਿੰਗ MapQTL ਵਿੱਚ ਅੰਤਰਾਲ ਮੈਪਿੰਗ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਕੀਤੀ ਗਈ ਸੀ27. MapQTL ਦੇ ਆਉਟਪੁੱਟ ਦੀ ਵਰਤੋਂ ਮੈਪਚਾਰਟ 2.3 ਨਾਲ QTL ਪਲਾਟ ਬਣਾਉਣ ਲਈ ਕੀਤੀ ਗਈ ਸੀ।28.
ਬਾਇਓਇਨਫੋਰਮੈਟਿਕ ਵਿਸ਼ਲੇਸ਼ਣ. ਸ਼ੁਰੂਆਤੀ 27.37 kb ਅੰਤਰਾਲ ਵਿੱਚ ਸਹੀ ਜੀਨ ਮਾਡਲਾਂ ਦੀ ਪਛਾਣ ਕਰਨ ਲਈ, ਅਸੀਂ DM4.04 ਸੰਦਰਭ ਜੀਨੋਮ, PGSC ਐਨੋਟੇਸ਼ਨ, ਅਤੇ ITAG ਐਨੋਟੇਸ਼ਨ ਲਈ ਦੋ ਵੱਖਰੀਆਂ ਜੀਨ ਐਨੋਟੇਸ਼ਨਾਂ ਦੀ ਜਾਂਚ ਕੀਤੀ। Hirsch et al ਨੂੰ ਵੀ ਦੇਖੋ।29. ਐਨੋਟੇਸ਼ਨਾਂ ਦੀ ਸ਼ੁੱਧਤਾ ਦੀ ਪੁਸ਼ਟੀ ਕਰਨ ਲਈ, ਅਸੀਂ ਦੋਵਾਂ ਐਨੋਟੇਸ਼ਨਾਂ ਤੋਂ ਅਨੁਮਾਨਿਤ ਪ੍ਰੋਟੀਨ ਕ੍ਰਮਾਂ ਦੇ ਨਾਲ BLASTp ਖੋਜਾਂ ਕੀਤੀਆਂ। ਸਾਡੀ ਪੁੱਛਗਿੱਛ ਨਾਲ BLASTp ਖੋਜ ਵਿੱਚ ਸਭ ਤੋਂ ਵਧੀਆ ਹਿੱਟ ਦੀ ਤੁਲਨਾ ਕਰਕੇ, ਅਸੀਂ ਇਹ ਨਿਰਧਾਰਿਤ ਕੀਤਾ ਕਿ ਕੀ ਭਵਿੱਖਬਾਣੀ ਪ੍ਰੋਟੀਨ ਕ੍ਰਮ ਵਿੱਚ ਸਾਰੇ ਐਨੋਟੇਟਿਡ ਐਕਸੋਨ ਅਤੇ ਡੋਮੇਨ ਆਲੂ ਅਤੇ ਹੋਰ ਪੌਦਿਆਂ ਦੀਆਂ ਕਿਸਮਾਂ ਵਿੱਚ ਸਮਾਨ ਪ੍ਰੋਟੀਨ ਦੁਆਰਾ ਸਮਰਥਿਤ ਸਨ ਜਾਂ ਨਹੀਂ। ਇਸ ਤੋਂ ਇਲਾਵਾ, SPUD DB 'ਤੇ ਜਨਤਕ ਤੌਰ 'ਤੇ ਉਪਲਬਧ RNA-seq ਲਾਇਬ੍ਰੇਰੀਆਂ (solanaceae.plantbiology.msu.edu/cgi-bin/ gbrowse/potato/ 'ਤੇ ਉਪਲਬਧ) ਅਤੇ NCBI ਜੀਨੋਮ ਡੇਟਾ ਵਿਊਅਰ (ncbi.nlm.nih.gov/ genome/gdv 'ਤੇ ਉਪਲਬਧ ਹੈ) /browser/) ਦੀ ਵਰਤੋਂ ਇਹ ਨਿਰਧਾਰਿਤ ਕਰਨ ਲਈ ਕੀਤੀ ਗਈ ਸੀ ਕਿ ਕੀ ਪੁਟੇਟਿਵ ਐਕਸੌਨ ਸਮੀਕਰਨ ਸਬੂਤ ਸਨ। ਇਕੱਠੇ ਮਿਲ ਕੇ, ਇਹਨਾਂ ਦੋ ਦ੍ਰਿਸ਼ਟੀਕੋਣਾਂ ਨੇ ਸਾਨੂੰ ਦੋਵਾਂ ਐਨੋਟੇਸ਼ਨਾਂ ਵਿੱਚ ਜੀਨ ਮਾਡਲਾਂ ਦੇ ਅੰਦਰੂਨੀ-ਐਕਸੋਨ ਢਾਂਚੇ ਨੂੰ ਪ੍ਰਮਾਣਿਤ ਕਰਨ ਦੀ ਇਜਾਜ਼ਤ ਦਿੱਤੀ, ਜਿਸਦੇ ਨਤੀਜੇ ਵਜੋਂ ਸਵਾਲ ਵਿੱਚ ਜੀਨ ਦੀ ਪ੍ਰਤੀਨਿਧਤਾ ਕਰਨ ਲਈ ਜੀਨ ਮਾਡਲਾਂ ਦੇ ਇੱਕ ਜਾਂ ਇੱਕ ਤੋਂ ਵੱਧ ਆਈਸੋਫਾਰਮ ਲਈ ਇੱਕ ਸੂਚਿਤ ਵਿਕਲਪ ਹੁੰਦਾ ਹੈ। ਇਹਨਾਂ ਪਹੁੰਚਾਂ ਦੇ ਆਧਾਰ 'ਤੇ, ਉਮੀਦਵਾਰ ਜੀਨ PGSC0003DMG400016862 ਨੂੰ ਸੰਭਾਵਤ ਤੌਰ 'ਤੇ ਅੰਸ਼ਕ ਅਤੇ ਮਾਮੂਲੀ ਤੌਰ 'ਤੇ ਪ੍ਰਗਟ ਕੀਤਾ ਗਿਆ ਸੀ ਅਤੇ ਹੋਰ ਵਿਸ਼ਲੇਸ਼ਣਾਂ ਤੋਂ ਰੱਦ ਕੀਤਾ ਗਿਆ ਸੀ। ਜੀਨ ਮਾਡਲ Sotub12g029970 ਨੂੰ ਸਹੀ ਮੰਨਿਆ ਗਿਆ ਸੀ, ਜਦੋਂ ਕਿ ਇਸਦਾ PGSC ਹਮਰੁਤਬਾ PGSC0003DMG400016860 ਸੰਭਾਵਤ ਤੌਰ 'ਤੇ ਕੱਟਿਆ ਗਿਆ ਹੈ। ਕਿਉਂਕਿ ਇਹ ਨਿਰਧਾਰਤ ਅੰਤਰਾਲ ਤੋਂ ਬਾਹਰ ਸਥਿਤ ਹੈ, ਅਤੇ SC ਅਤੇ SI ਪੌਦਿਆਂ ਦੇ ਵਿਚਕਾਰ ਕੋਈ ਵੀ ਸੰਬੰਧਿਤ ਅਮੀਨੋ ਐਸਿਡ ਬਦਲਾਂ ਦੀ ਪਛਾਣ ਨਹੀਂ ਕੀਤੀ ਜਾ ਸਕਦੀ ਹੈ, ਇਸ ਜੀਨ ਨੂੰ ਹੋਰ ਵਿਸ਼ਲੇਸ਼ਣਾਂ ਤੋਂ ਖਾਰਜ ਕਰ ਦਿੱਤਾ ਗਿਆ ਸੀ।
ਪਰਿਵਰਤਨ ਵਿਸ਼ਲੇਸ਼ਣ. 27.37 kb ਅੰਤਰਾਲ ਵਿੱਚ ਪਰਿਵਰਤਨ ਦੀ ਪਛਾਣ ਕਰਨ ਲਈ ਜੋ ਸਵੈ-ਅਨੁਕੂਲ ਜੀਨੋਟਾਈਪਾਂ ਲਈ ਖਾਸ ਹਨ, ਸਾਰੇ ਉੱਚ ਆਤਮ ਵਿਸ਼ਵਾਸ ਵਾਲੇ SNPs (ਪੂਰਕ ਡੇਟਾ 3) ਨਿਰਧਾਰਤ ਕੀਤੇ ਗਏ ਸਨ ਜੋ ਕਿ (1) DS, 17SC100-18 ਅਤੇ 17SC100-2 ਵਿੱਚ ਸਮਰੂਪ ਸਨ (ਕਿਉਂਕਿ ਤਿੰਨੋਂ ਹੀ ਹੋਮੋਜ਼ਾਈਗਸ ਲਈ ਹਨ। ਦਾ SC ਐਲੀਲ ਸਲੀ (Sli/Sli')'), (2) D16 ਵਿੱਚ ਸਮਰੂਪਿਕ ਵੱਖਰਾ ਹੈ (ਕਿਉਂਕਿ D16 Sli (sli/sli) ਦੇ SI ਐਲੀਲ ਲਈ ਸਮਰੂਪ ਹੈ), ਅਤੇ (3) 16HP1-66 ਅਤੇ 17SC25-8 ਦੋਵਾਂ ਵਿੱਚ ਵਿਪਰੀਤ ਜ਼ਾਇਗਸ ਹੈ (ਕਿਉਂਕਿ ਦੋਵੇਂ SC ਐਲੀਲ Sli (Sli) ਲਈ ਵਿਪਰੀਤ ਹਨ। /sli)). ਐਲੇਲਿਕ ਕ੍ਰਮ SPAdes ਸੰਸਕਰਣ 3.11.1 ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਡੀ ਨੋਵੋ ਅਸੈਂਬਲੀ ਦੁਆਰਾ ਪ੍ਰਾਪਤ ਕੀਤਾ ਗਿਆ ਸੀ30 ਉੱਪਰ-ਸੂਚੀਬੱਧ ਪੌਦਿਆਂ ਦੇ 150 nt ਪੇਅਰਡ-ਐਂਡ ਇਲੂਮਿਨਾ ਡੇਟਾ (ਲਗਭਗ 25-30X ਕ੍ਰਮ ਦੀ ਡੂੰਘਾਈ ਦਾ)। ਨਤੀਜੇ ਵਾਲੇ ਕੰਟਿਗਸ ਨੂੰ DM ਸੰਦਰਭ (minimap2 ਸੰਸਕਰਣ 2.1 ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ) ਨਾਲ ਇਕਸਾਰ ਕੀਤਾ ਗਿਆ ਸੀ ਅਤੇ ਉਹਨਾਂ ਲਈ ਫਿਲਟਰ ਕੀਤਾ ਗਿਆ ਸੀ ਜੋ 27 kb ਨਾਲ ਭਰੋਸੇਯੋਗ ਤੌਰ 'ਤੇ ਅਲਾਈਨ ਹੁੰਦੇ ਹਨ। ਇਹਨਾਂ ਅਲਾਈਨਡ ਕੰਟਿਗਸ ਤੋਂ, DM4.03 ਦੇ ਅਨੁਸਾਰੀ ਪਰਿਵਰਤਨ ਨੂੰ ਸਿੱਧੇ ਤੌਰ 'ਤੇ ਮਾਪਿਆ ਗਿਆ ਸੀ (subroutines mpileup ਅਤੇ bcftools, ਸੰਸਕਰਣ 1.9 ਤੋਂ ਕਾਲ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ) ਅਤੇ ਵੇਰੀਐਂਟ ਕਾਲ ਫਾਰਮੈਟ (VCF) ਵਿੱਚ ਸੂਚੀਬੱਧ ਕੀਤਾ ਗਿਆ ਸੀ।
ਅਮੀਨੋ ਐਸਿਡ ਤਬਦੀਲੀ ਦਾ ਵਿਸ਼ਲੇਸ਼ਣ. SC-ਵਿਸ਼ੇਸ਼ ਪਰਿਵਰਤਨ ਦੀ ਇਸ ਸੂਚੀ ਤੋਂ, ਸਾਰੇ ਗੈਰ-ਸਮਾਨਯੋਗ SNPs ਨੂੰ ਮਨੋਨੀਤ ਕੋਡਿੰਗ ਐਕਸੌਨ ਨਾਲ ਓਵਰਲੈਪ ਕਰਕੇ ਪਛਾਣਿਆ ਗਿਆ ਸੀ। DM ਜਾਂ SI ਕ੍ਰਮ ਦੇ ਅਨੁਸਾਰੀ ਅਮੀਨੋ ਐਸਿਡ ਤਬਦੀਲੀਆਂ ਨੂੰ ਸੂਚੀਬੱਧ ਕੀਤਾ ਗਿਆ ਸੀ। ਪ੍ਰੋਟੀਨ ਕ੍ਰਮ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ BLASTp ਖੋਜਾਂ ਕਰਨ ਅਤੇ ਚੋਟੀ ਦੇ 100 BLASTp ਹਿੱਟਾਂ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਮਲਟੀਪਲ ਕ੍ਰਮ ਅਲਾਈਨਮੈਂਟ ਕਰਨ ਦੁਆਰਾ ਵਿਲੱਖਣ ਅਮੀਨੋ ਐਸਿਡ ਤਬਦੀਲੀਆਂ ਦੀ ਪਛਾਣ ਕੀਤੀ ਗਈ ਸੀ।
ਪ੍ਰਮੋਟਰ ਅਤੇ ਟਰਮੀਨੇਟਰ ਖੇਤਰਾਂ ਵਿੱਚ ਪਰਿਵਰਤਨ. ਪ੍ਰਮੋਟਰ ਖੇਤਰ ਨੂੰ ਸਟਾਰਟ ਕੋਡਨ ਦਾ ਕ੍ਰਮ ਅੱਪਸਟਰੀਮ ਹੋਣ ਲਈ ਚੁਣਿਆ ਗਿਆ ਸੀ ਜਦੋਂ ਤੱਕ ਕਿ ਵੱਧ ਤੋਂ ਵੱਧ 1500 nt ਦੇ ਨਾਲ ਅੱਪਸਟਰੀਮ ਜੀਨ ਦੇ ਕੋਡਿੰਗ ਕ੍ਰਮ ਤੱਕ. ਪ੍ਰਮੋਟਰ ਖੇਤਰਾਂ ਵਿੱਚ ਨਾਟਕੀ ਪਰਿਵਰਤਨ 27.37 kb ਅੰਤਰਾਲ ਦੇ ਅੰਦਰ ਪਾਇਆ ਗਿਆ ਸੀ, ਜਿਸ ਵਿੱਚ ਸਭ ਤੋਂ ਪ੍ਰਭਾਵਸ਼ਾਲੀ ਕਈ ਵੱਡੇ ਮਿਟਾਏ ਗਏ ਸਨ ਅਤੇ ਦਸਾਂ ਤੋਂ ਸੈਂਕੜੇ ਨਿਊਕਲੀਓਟਾਈਡ ਲੰਬਾਈ ਦੇ ਸੰਮਿਲਨ ਸਨ। Sli ਅੰਤਰਾਲ ਵਿੱਚ ਸਾਰੇ ਪਰਿਵਰਤਨ, DM ਦੇ ਅਨੁਸਾਰੀ, ਪ੍ਰਾਪਤ ਕੀਤੇ ਗਏ ਸਨ, ਜਿਸ ਵਿੱਚ ਪ੍ਰਮੋਟਰ/ਅੱਪਸਟ੍ਰੀਮ ਖੇਤਰ ਦੇ ਨਾਲ-ਨਾਲ ਟਰਮੀਨੇਟਰ/ਡਾਊਨਸਟ੍ਰੀਮ ਖੇਤਰ ਵੀ ਸ਼ਾਮਲ ਹੈ।
ਪਰਾਗ ਪ੍ਰਾਪਤੀ ਅਤੇ ਉਗਣ. ਚਿੱਤਰ ਵਿੱਚ ਸੂਚੀਬੱਧ ਜੀਨੋਟਾਈਪਾਂ ਤੋਂ ਪਰਾਗ। 2a ਨੂੰ ਇਲੈਕਟ੍ਰਾਨਿਕ ਟੂਥਬ੍ਰਸ਼ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਖੁੱਲੇ ਫੁੱਲਾਂ ਨੂੰ ਵਾਈਬ੍ਰੇਟ ਕਰਕੇ ਅਤੇ 1.5 ਮਿਲੀਲੀਟਰ ਐਪੇਨਡੋਰਫ ਟਿਊਬਾਂ ਵਿੱਚ ਪਰਾਗ ਨੂੰ ਇਕੱਠਾ ਕਰਕੇ ਪ੍ਰਾਪਤ ਕੀਤਾ ਗਿਆ ਸੀ। ਗ੍ਰਹਿਣ ਕਰਨ ਤੋਂ ਬਾਅਦ, ਪਰਾਗ ਨੂੰ ਕਮਰੇ ਦੇ ਤਾਪਮਾਨ 'ਤੇ 24 ਘੰਟਿਆਂ ਲਈ ਸਿਲਿਕਾ ਜੈੱਲ ਵਾਲੇ ਹਵਾ-ਸੀਲਬੰਦ ਬਕਸੇ ਵਿੱਚ ਪਰਾਗ ਦੇ ਨਾਲ ਖੁੱਲ੍ਹੀਆਂ ਐਪੇਨਡੋਰਫ ਟਿਊਬਾਂ ਨੂੰ ਸਟੋਰ ਕਰਕੇ ਸੁਕਾਇਆ ਗਿਆ ਸੀ। ਬਾਅਦ ਵਿੱਚ, ਪਰਾਗ ਨੂੰ ਅਗਲੀ ਵਰਤੋਂ ਤੱਕ -20 ° C 'ਤੇ ਸਟੋਰ ਕੀਤਾ ਗਿਆ ਸੀ।
ਪਰਾਗ ਨੂੰ 2.5 ਮਿਲੀਲੀਟਰ ਤਰਲ ਮਾਧਿਅਮ (5% (w/v) ਸੁਕਰੋਜ਼, 9 mg/l ਬੋਰਿਕ ਐਸਿਡ, 50 mg/l CaCly73.5H ਵਿੱਚ 2 ਮਿਲੀਗ੍ਰਾਮ ਸੁੱਕੇ ਪਰਾਗ ਨੂੰ ਮੁਅੱਤਲ ਕਰਕੇ ਉਗਾਇਆ ਗਿਆ ਸੀ।2O, 118 mg/l Ca (NO3) 24 ਐੱਚ2O, 123 mg/l MgSO.4VH2O) ਪੈਰਾਫਿਲਮ ਨਾਲ ਸੀਲ ਕੀਤੇ 3.5 ਸੈਂਟੀਮੀਟਰ ਵਿਆਸ ਵਾਲੇ ਪੈਟਰੀ ਪਕਵਾਨਾਂ ਵਿੱਚ। ਪਰਾਗ ਨੂੰ ਕਮਰੇ ਦੇ ਤਾਪਮਾਨ 'ਤੇ ਹਿੱਲਦੇ ਹੋਏ ਅਤੇ 24 RPM 'ਤੇ ਹਿੱਲਣ ਵਾਲੇ ਇਨਕਿਊਬੇਟਰ ਵਿੱਚ ਹਨੇਰੇ ਵਿੱਚ 125 ਘੰਟਿਆਂ ਲਈ ਪੈਟਰੀ ਡਿਸ਼ਾਂ ਵਿੱਚ ਉਗਣ ਲਈ ਛੱਡ ਦਿੱਤਾ ਗਿਆ ਸੀ। ਉਗਣ ਵਾਲੇ ਪਰਾਗ ਵਾਲੇ ਤਰਲ ਮਾਧਿਅਮ ਨੂੰ ਫਿਰ ਧਿਆਨ ਨਾਲ ਪਾਈਪੇਟ ਟਿਪਸ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ 2 ਮਿਲੀਲੀਟਰ ਏਪੇਨਡੋਰਫ ਟਿਊਬਾਂ ਵਿੱਚ ਪਾਈਪ ਕੀਤਾ ਗਿਆ ਸੀ ਜੋ ਅਪਰਚਰ ਦਾ ਆਕਾਰ ਵਧਾਉਣ ਲਈ ਸੋਧਿਆ ਗਿਆ ਸੀ ਤਾਂ ਜੋ ਪਰਾਗ ਟਿਊਬਾਂ ਨੂੰ ਨੁਕਸਾਨ ਨਾ ਪਹੁੰਚ ਸਕੇ। Eppendorf ਟਿਊਬਾਂ ਨੂੰ ਫਿਰ 600 ਮਿੰਟ ਲਈ 1xg 'ਤੇ ਸੈਂਟਰਿਫਿਊਜ ਕੀਤਾ ਗਿਆ ਸੀ ਅਤੇ ਮਾਧਿਅਮ ਨੂੰ ਪਾਈਪਟਿੰਗ ਦੁਆਰਾ ਧਿਆਨ ਨਾਲ ਹਟਾ ਦਿੱਤਾ ਗਿਆ ਸੀ। ਪੈਲੇਟ ਅਤੇ ਕੁਝ ਬਾਕੀ ਬਚੇ ਮਾਧਿਅਮ ਨੂੰ ਤੁਰੰਤ ਤਰਲ ਨਾਈਟ੍ਰੋਜਨ ਵਿੱਚ ਫ੍ਰੀਜ਼ ਕੀਤਾ ਗਿਆ ਸੀ, ਦੋ ਸਟੇਨਲੈਸ ਸਟੀਲ ਮਣਕੇ (2 ਮਿਲੀਮੀਟਰ ਵਿਆਸ) ਜੋੜੇ ਗਏ ਸਨ ਅਤੇ ਨਮੂਨਿਆਂ ਨੂੰ 20 ਮਿੰਟ ਲਈ 1 Hz 'ਤੇ ਟਿਸ਼ੂਲਾਈਜ਼ਰ II (ਕਿਆਗੇਨ GmbH, ਹਿਲਡਨ, ਜਰਮਨੀ) ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਪੀਸਿਆ ਗਿਆ ਸੀ।
ਆਰਐਨਏ ਕੱਢਣ ਅਤੇ ਕ੍ਰਮਬੱਧ. ਬਫਰ RLT (Qiagen GmbH) ਨੂੰ ਪੀਸ ਕੀਤੇ ਪਰਾਗ ਦੇ ਨਮੂਨਿਆਂ ਵਿੱਚ ਜੋੜਿਆ ਗਿਆ ਜਦੋਂ ਕਿ ਇਹ ਯਕੀਨੀ ਬਣਾਇਆ ਗਿਆ ਕਿ ਨਮੂਨੇ ਜੰਮੇ ਰਹੇ। ਆਰਐਨਏ ਐਕਸਟਰੈਕਸ਼ਨ ਫਿਰ ਨਿਰਮਾਤਾ ਦੇ ਪ੍ਰੋਟੋਕੋਲ (ਕਿਆਗੇਨ ਜੀਐਮਬੀਐਚ, ਹਿਲਡਨ, ਜਰਮਨੀ) ਦੇ ਅਨੁਸਾਰ ਆਰਨਸੀ ਮਿੰਨੀ ਕਿੱਟ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਕੀਤੀ ਗਈ ਸੀ। 250-300 bp ਇਨਸਰਟ-ਸਾਈਜ਼ cDNA ਲਾਇਬ੍ਰੇਰੀਆਂ ਨੂੰ 150nt ਪੇਅਰਡ-ਐਂਡ ਰੀਡਜ਼ ਵਜੋਂ ਕ੍ਰਮਬੱਧ ਕੀਤਾ ਗਿਆ ਸੀ, ਪ੍ਰਤੀ ਨਮੂਨਾ 30-42 ਮਿਲੀਅਨ ਰੀਡ-ਪੇਅਰਸ (ਨੋਵੋਜੀਨ, ਕੈਮਬ੍ਰਿਜ, ਯੂਨਾਈਟਿਡ ਕਿੰਗਡਮ)।
ਹੋਰ RNA-seq ਡਾਟਾ ਸੈੱਟ. (ਟਿਸ਼ੂ-ਵਿਸ਼ੇਸ਼) ਸਮੀਕਰਨ ਪੱਧਰਾਂ ਦੀ ਸੰਖੇਪ ਜਾਣਕਾਰੀ ਬਣਾਉਣ ਲਈ, ਸਾਰੇ ਪੇਅਰਡ-ਐਂਡ ਕ੍ਰਮਬੱਧ RNA-seq ਡੇਟਾ ਸੈੱਟਾਂ ਨੂੰ ORGANISM ਵਜੋਂ ਟੈਗ ਕੀਤਾ ਗਿਆ ਹੈ "ਸੋਲਾ-ਨਮ ਟਿਊਬਰੋਸਮ" ਜਨਤਕ ਡੋਮੇਨ (NCBI-SRA, ਮਿਤੀ 2018/17/13) ਤੋਂ ਡਾਊਨਲੋਡ ਕੀਤੇ ਗਏ ਸਨ, ਕੁੱਲ 441 ਪੇਅਰ ਕੀਤੇ fastq ਡਾਟਾ ਸੈੱਟ। ਇਹਨਾਂ 441 ਜਨਤਕ ਡੇਟਾ ਸੈੱਟਾਂ ਵਿੱਚੋਂ, 3 ਸਟਾਈਲ ਟਿਸ਼ੂ (SRR7402817-SRR7402819) ਤੋਂ ਅਤੇ ਬਾਕੀ ਸਾਰੇ ਵੱਖ-ਵੱਖ ਗੈਰ-ਪਰਾਗ ਟਿਸ਼ੂਆਂ, ਵਿਕਾਸ ਦੇ ਪੜਾਵਾਂ, ਅਤੇ ਪੌਦਿਆਂ ਦੇ ਐਕਸੈਸਸ਼ਨ ਤੋਂ ਤਿਆਰ ਕੀਤੇ ਗਏ ਸਨ।
Solyntus ਸੰਦਰਭ ਅਸੈਂਬਲੀ. ਸਮੀਕਰਨ ਵਿਸ਼ਲੇਸ਼ਣ ਲਈ, ਸਮਰੂਪ ਸੰਦਰਭ ਲਾਈਨ ਸੋਲਿਨਟਸ (ਵਰਜਨ 1.0, ਇੱਥੇ ਡਾਊਨਲੋਡ ਕਰਨਯੋਗ) ਦੀ ਹਾਲ ਹੀ ਵਿੱਚ ਪ੍ਰਾਪਤ ਕੀਤੀ ਡਰਾਫਟ ਅਸੈਂਬਲੀ www.plantbreeding.wur.nl/Solyntus/) ਨੂੰ ਇੱਕ ਹਵਾਲਾ ਜੀਨੋਮ ਵਜੋਂ ਵਰਤਿਆ ਗਿਆ ਸੀ। ਸੋਲਿੰਟਾ ਦੇ ਪ੍ਰਜਨਨ ਪ੍ਰੋਗਰਾਮ ਦੇ ਹਿੱਸੇ ਵਜੋਂ ਤਿਆਰ ਕੀਤੀ ਗਈ ਇੱਕ ਜ਼ਰੂਰੀ ਸਮਰੂਪ ਕਿਸਮ ਹੈ।21. ਇਸ ਅਧਿਐਨ ਵਿੱਚ ਮੈਪਿੰਗ ਅੰਤਰਾਲਾਂ ਦਾ DM v4.03 ਜੀਨੋਮ ਅਸੈਂਬਲੀ ਤੋਂ ਅਨੁਮਾਨ ਲਗਾਇਆ ਗਿਆ ਸੀ31 (Solyntus 1.0 ਜੀਨੋਮ ਅਸੈਂਬਲੀ ਕੋਆਰਡੀਨੇਟਸ) 1.0-53532708 (ਅੰਤਰਾਲ I, 53954293kb <—421.6kb), 628.9kb <—53683239kb, 53867377-184.1 'ਤੇ ਸਥਿਤ ਹੋਣ ਲਈ ਬੁਨਿਆਦੀ ਸਮਾਨਤਾ ਖੋਜਾਂ (BLASTn ਅਤੇ ਬੈੱਡਟੂਲਸ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ) ਦੁਆਰਾ Solyntus 168.7 ਜੀਨੋਮ ਅਸੈਂਬਲੀ ਲਈ kb< — 53731620kb), 53763003-31.4 (ਅੰਤਰਾਲ III, 27.4kb< —53753977kb) ਅਤੇ 53763003-9.0 (ਅੰਤਰਾਲ IV, 12.6kb < —1.0 kb), ਕ੍ਰਮਵਾਰ। ਬਰੈਕਟਾਂ ਦੇ ਵਿਚਕਾਰ ਕ੍ਰਮਵਾਰ ਲਗਾਤਾਰ ਮੈਪਿੰਗ ਅੰਤਰਾਲ ਨੰਬਰ [ਸੋਲਿਨਟਸ 1.0 ਕੋਆਰਡੀਨੇਟਸ], ਸੋਲਿੰਟਸ-4.03 ਵਿੱਚ ਆਕਾਰ ਅਤੇ DM-4.04/4.03 ਵਿੱਚ ਆਕਾਰ ਹਨ। ਸਾਰੇ ਅੰਤਰਾਲ ਕ੍ਰੋਮੋਸੋਮ ST12ch12_RaGOO (ਕ੍ਰੋਮੋਸੋਮ 1.0 ਹੋਣ) 'ਤੇ ਸਥਿਤ ਹੁੰਦੇ ਹਨ ਅਤੇ ਸੋਲਿਨਟਸ 4.03 ਅਸੈਂਬਲੀ ਵਿੱਚ ਇੱਕ ਵੀ ਅੰਤਰ ਨਹੀਂ ਹੁੰਦਾ। ਅੰਤਰਾਲ ਆਕਾਰ ਪਰਿਵਰਤਨ ਸੰਬੰਧਿਤ DM ਕ੍ਰਮ ਵਿੱਚ ਬਹੁਤ ਸਾਰੇ ਅੰਤਰ (N's) ਅਤੇ ਦੋਵਾਂ ਜੀਨੋਮ ਦੇ ਵਿਚਕਾਰ ਵਿਆਪਕ ਪਰਿਵਰਤਨ ਦੇ ਕਾਰਨ ਹੁੰਦਾ ਹੈ। DM ਜੀਨੋਮ (DM-4.04/12) 'ਤੇ ਅਨੁਸਾਰੀ ਅੰਤਰਾਲ: ਅੰਤਰਾਲ I: chr58601503:59230363-12, ਅੰਤਰਾਲ II: chr58962004:59130723-59016142; ਅੰਤਰਾਲ III: 59043512-12; ਅੰਤਰਾਲ IV: chr59030880:59043512-XNUMX।
Solyntus 1.0 'ਤੇ ਜੀਨ ਐਨੋਟੇਸ਼ਨ ਦਾ ਅਨੁਮਾਨ ਤਿੰਨ ਵੱਖ-ਵੱਖ ਜੀਨ ਕੈਟਾਲਾਗਾਂ (ਆਲੂ DM4.03, ITAG4.0 ਟਮਾਟਰ ਜੀਨੋਮ ਐਨੋਟੇਸ਼ਨ 6 ਸਤੰਬਰ, 2019 ਦੀ ਰਿਲੀਜ਼) ਤੋਂ ਲਗਾਇਆ ਗਿਆ ਸੀ।32, ਅਤੇ Pepper-v. 1.5533), ਜੋ GeMoMa (v1.6.1) ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਸੋਲੀਟਸ ਅਸੈਂਬਲੀ ਵਿੱਚ ਮੈਪ ਕੀਤੇ ਗਏ ਸਨ। ਇਹ ਵਿਅਕਤੀਗਤ ਜੀਨ ਕੈਟਾਲਾਗ ਵਿੱਚ ਕਮੀਆਂ ਦੀ ਪੂਰਤੀ ਲਈ ਅਤੇ ਸੰਭਵ ਜੀਨਾਂ ਅਤੇ/ਜਾਂ ਪ੍ਰਗਟ ਕੀਤੇ ਸਥਾਨਾਂ ਦੀ ਮੌਜੂਦਗੀ ਬਾਰੇ ਸਾਡੀ ਜਾਗਰੂਕਤਾ ਨੂੰ ਵੱਧ ਤੋਂ ਵੱਧ ਕਰਨ ਲਈ ਕੀਤਾ ਗਿਆ ਸੀ।
RNA-seq ਰੀਡ-ਮੈਪਿੰਗ ਅਤੇ ਟ੍ਰਾਂਸਕ੍ਰਿਪਟ ਭਰਪੂਰ ਮਾਤਰਾfiਕੇਟੇਸ਼ਨ. ਸਾਰੇ 5 SC, 3 SI, ਅਤੇ ਸਾਰੇ 441 ਜਨਤਕ RNA-seq ਡੇਟਾ ਸੈੱਟਾਂ ਨੂੰ hisat2 (ਵਰਜਨ 2.1.0) ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਸੋਲਿਨਟਸ ਸੰਦਰਭ ਜੀਨੋਮ ਨਾਲ ਮੈਪ ਕੀਤਾ ਗਿਆ ਸੀ। GeMoMa ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਪ੍ਰਾਪਤ ਕੀਤੇ ਗਏ ਹਾਈਬ੍ਰਿਡ ਜੀਨ ਕੈਟਾਲਾਗ ਦੀ ਵਰਤੋਂ ਸਟ੍ਰਿੰਗਟਾਈ (ਵਰਜਨ 2.1.1) ਸੈਟਿੰਗਾਂ -t -c 5 -f 0.05 -G ਅਤੇ ਇੱਕ GeMoMa ਸੰਯੋਜਿਤ Solyntus1.0 gff ਫਾਈਲ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਟ੍ਰਾਂਸਕ੍ਰਿਪਟ-ਗਾਈਡਿਡ ਭਰਪੂਰਤਾ ਅਨੁਮਾਨ ਲਈ ਕੀਤੀ ਗਈ ਸੀ। ਸਾਰੇ ਦੇ ਆਲੇ-ਦੁਆਲੇ 500 kb ਅੰਤਰਾਲ ਵਿੱਚ ਸਮੀਕਰਨ ਦੇਖਿਆ ਗਿਆ ਸਲੀ ਇੱਕ ਕੇਂਦਰ ਵਜੋਂ ਸਥਾਨ ਦਾ ਮੁਲਾਂਕਣ ਕੀਤਾ ਗਿਆ ਸੀ, ਜਿਸ ਵਿੱਚ ਅੰਤਰਾਲ ਵਿੱਚ ਕੁੱਲ 90 (ਅਨੁਮਾਨਿਤ) ਜੀਨ ਲੋਕੀ ਸਥਿਤ ਹਨ। ਅਸੀਂ ਇਹਨਾਂ ਵਿੱਚੋਂ ਕਿਸੇ ਵੀ ਜੀਨ ਸਥਾਨ ਦੇ ਬਾਹਰ SC ਨਮੂਨਿਆਂ ਵਿੱਚ ਕਿਸੇ ਵੀ ਧਿਆਨ ਦੇਣ ਯੋਗ ਸਮੀਕਰਨ ਦੀ ਅਣਹੋਂਦ ਦੀ ਪੁਸ਼ਟੀ ਕੀਤੀ ਹੈ। 500 kb ਅੰਤਰਾਲ ਵਿੱਚ, ਅਸੀਂ ਉੱਪਰ ਪਰਿਭਾਸ਼ਿਤ ਕੀਤੇ ਅਨੁਸਾਰ ਸਾਡੇ ਮੈਪਿੰਗ ਅੰਤਰਾਲ I-IV ਦੇ ਨਾਲ ਕੱਟਦੇ ਹੋਏ ਉਮੀਦਵਾਰਾਂ ਦੇ ਜੀਨਾਂ ਦੀ ਬਾਅਦ ਵਿੱਚ ਛੋਟੀ ਸੰਖਿਆ ਦਾ ਸੰਕੇਤ ਦਿੱਤਾ।
cuntfiਹੈਪਲੋਟਾਈਪ-ਵਿਸ਼ੇਸ਼ਤਾ ਦੀ ਰਚਨਾfic ਸਮੀਕਰਨ. 90 kb ਅੰਤਰਾਲ ਵਿੱਚ 500 ਪ੍ਰਗਟ ਕੀਤੇ ਸਥਾਨਾਂ ਵਿੱਚੋਂ, ਸਾਰੇ SC/SI ਨਮੂਨਿਆਂ ਵਿੱਚ 8 FPKM ਦੀ ਇੱਕ ਚੁਣੀ ਹੋਈ ਥ੍ਰੈਸ਼ਹੋਲਡ ਤੋਂ ਉੱਪਰ ਸਿਰਫ਼ 20 ਨੂੰ ਦਰਸਾਇਆ ਗਿਆ ਸੀ। ਅਸੀਂ ਇਹਨਾਂ ਸਾਈਟਾਂ ਦੀ ਵਰਤੋਂ ਹੈਪਲੋਟਾਈਪ-ਵਿਸ਼ੇਸ਼ (Sli ਜਾਂ sli) ਸਮੀਕਰਨ ਪੱਧਰ ਦੇ ਅੰਤਰ ਨੂੰ ਮਾਪਣ ਲਈ ਕੀਤੀ ਹੈ। ਚੁਣੀ ਗਈ ਸਮੀਕਰਨ ਥ੍ਰੈਸ਼ਹੋਲਡ ਨੇ ਅੰਤ ਵਿੱਚ ਸਮੀਕਰਨ ਨੂੰ (ਵੱਧ ਤੋਂ ਵੱਧ) 2 ਹੈਪਲੋਟਾਈਪਾਂ ਵਿੱਚ ਭਰੋਸੇਮੰਦ ਢੰਗ ਨਾਲ ਪੜਾਅ ਕਰਨ ਲਈ ਕਾਫ਼ੀ ਪੜ੍ਹਨ ਦੀ ਡੂੰਘਾਈ ਨੂੰ ਸਮਰੱਥ ਬਣਾਇਆ। ਖੁਦ PSC ਲੋਕਸ (ਜਿਸ ਵਿੱਚ SI ਪੌਦਿਆਂ ਵਿੱਚ ਪ੍ਰਗਟਾਵੇ ਦੀ ਘਾਟ ਹੈ) ਦੇ ਨਾਲ, ਇਹ 8 + 1 ਲੋਕੀ 8 ਨਮੂਨਿਆਂ ਵਿੱਚੋਂ ਹਰੇਕ ਵਿੱਚ ਹੈਪਲੋਟਾਈਪ ਕੀਤੇ ਗਏ ਸਨ (SAMtools ਪੜਾਅ ਸੰਸਕਰਣ 1.7, ਡਿਫੌਲਟ ਸੈਟਿੰਗਜ਼)। ਨਤੀਜੇ ਵਜੋਂ ਹੈਪਲੋਟਾਈਪਡ (ਪੇਅਰਡ) ਫਾਸਟਕ ਫਾਈਲਾਂ ਨੂੰ SPAdes (ਵਰਜਨ 3.11.1) ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਡੀ ਨੋਵੋ ਅਸੈਂਬਲ ਕੀਤਾ ਗਿਆ ਸੀ। ਨਤੀਜੇ ਵਜੋਂ ਤਿਆਰ ਕੀਤੇ ਗਏ ਕੰਟਿਗਜ਼ ਨੂੰ ਭਰਪੂਰ ਮਾਤਰਾ ਲਈ ਫਿਲਟਰ ਕੀਤਾ ਗਿਆ ਸੀ ਅਤੇ ਮੁੱਖ (ਹੈਪਲੋਟਾਈਪਡ) ਪ੍ਰਗਟ ਕੀਤੇ ਆਈਸੋਫਾਰਮ ਦੇ ਅਨੁਸਾਰੀ ਪੂਰੀ-ਲੰਬਾਈ ਵਾਲੇ mRNAs ਦਾ ਅਨੁਮਾਨ ਲਗਾਇਆ ਗਿਆ ਸੀ। ਕੁਝ ਮਾਮਲਿਆਂ ਵਿੱਚ, ਇਸ ਨੇ ਵਿਕਲਪਿਕ ਤੌਰ 'ਤੇ ਕੱਟੇ ਹੋਏ ਆਈਸੋਫਾਰਮਾਂ ਨੂੰ ਹਟਾ ਦਿੱਤਾ, ਜਿਨ੍ਹਾਂ ਵਿੱਚੋਂ ਕੋਈ ਵੀ ਸਪੱਸ਼ਟ ਜੈਵਿਕ ਮਹੱਤਤਾ ਦੇ ਹੋਣ ਲਈ ਕਾਫ਼ੀ ਰੀਡ ਦੁਆਰਾ ਸਮਰਥਿਤ ਨਹੀਂ ਸੀ। ਇਹਨਾਂ ਹੈਪਲੋਟਾਈਪਡ mRNA ਕ੍ਰਮਾਂ ਵਿੱਚ ਪਰਿਵਰਤਨ ਦੀ ਵਰਤੋਂ (ਡਿਸ) ਕਰਨ ਲਈ ਕੀਤੀ ਗਈ ਸੀ ਕਿ ਕੀ ਇੱਕ ਜਾਂ ਦੋਵੇਂ ਹੈਪਲੋਟਾਈਪ ਹਰੇਕ ਸੰਬੰਧਿਤ ਸਥਾਨ/ਨਮੂਨਿਆਂ ਵਿੱਚ ਪ੍ਰਗਟ ਕੀਤੇ ਗਏ ਸਨ।
ਦੀ ਡਿਜ਼ਾਈਨ ਸਲੀ ਸਮੀਕਰਨ ਰਚਨਾ. ਅਸੀਂ Sli ਸਮੀਕਰਨ ਕੈਸੇਟ ਨੂੰ ਡਿਜ਼ਾਈਨ ਕਰਨ ਲਈ Sli ਡੋਨਰ ਪਲਾਂਟ DS ਦੇ ਕ੍ਰਮ ਦੀ ਵਰਤੋਂ ਕੀਤੀ। ਦੇ ਮੂਲ ਪ੍ਰਗਟਾਵੇ ਦੀ ਆਗਿਆ ਦੇਣ ਲਈ PSC, ਅਸੀਂ ਨੇਟਿਵ ਪ੍ਰਮੋਟਰ (ਸਟਾਰਟ ਕੋਡਨ ਦਾ 1563 bp ਅੱਪਸਟ੍ਰੀਮ), ਤਿੰਨ ਐਕਸੌਨ, ਅਤੇ ਨੇਟਿਵ ਟਰਮੀਨੇਟਰ (ਸਟੌਪ ਕੋਡਨ ਦਾ 740 bp ਡਾਊਨਸਟ੍ਰੀਮ) ਸਮੇਤ ਇੱਕ ਨਿਊਕਲੀਕ ਐਸਿਡ ਕ੍ਰਮ ਦਾ ਨਿਰਮਾਣ ਕੀਤਾ। ਇਸ ਪ੍ਰਕਾਰ, ਦੋਨੋਂ ਇੰਦਰਾਜ਼ਾਂ ਨੂੰ ਹਟਾ ਦਿੱਤਾ ਗਿਆ ਸੀ ਪੀ ਐੱਸ ਸੀ ਦਾਨੀ ਪੌਦੇ DS ਦਾ ਜੀਨ. ਇਸ ਕ੍ਰਮ ਨੂੰ ਜੈਨਸਕਰਿਪਟ (ਜੇਨਸਕਰਿਪਟ ਬਾਇਓਟੈਕ, ਲੀਡੇਨ, ਨੀਦਰਲੈਂਡਜ਼) ਦੁਆਰਾ ਸੰਸ਼ਲੇਸ਼ਣ ਅਤੇ ਕਲੋਨ ਕੀਤਾ ਗਿਆ ਸੀ। ਅਸੀਂ ਵੈਕਟਰ ਦਾ ਹਵਾਲਾ ਦਿੰਦੇ ਹਾਂ ਜਿਸ ਵਿੱਚ ਸਲੀ pBINPLUS-Sli ਵਜੋਂ ਸੰਮਿਲਿਤ ਕਰੋ।
CRISPR ਦਾ ਨਿਰਮਾਣ-Cas9 ਵੈਕਟਰ. ਅਸੀਂ DM0003 ਵਿੱਚ PGSC400016861DMG4.03 ਦੇ ਕ੍ਰਮ ਦੇ ਆਧਾਰ 'ਤੇ ਚਾਰ ਜੀਆਰਐਨਏ ਡਿਜ਼ਾਈਨ ਕੀਤੇ ਹਨ ਜਿੱਥੇ SC ਅਤੇ SI ਐਲੀਲਾਂ ਵਿਚਕਾਰ ਕੋਈ ਭਿੰਨਤਾ ਮੌਜੂਦ ਨਹੀਂ ਸੀ। ਢੁਕਵੇਂ ਗਾਈਡਾਂ ਦੀ ਚੋਣ ਅਤੇ ਵੈਕਟਰ ਦੀ ਉਸਾਰੀ ਲਈ, ਅਸੀਂ ਸੈਂਟੀਲਨ ਮਾਰਟੀਨੇਜ਼ ਐਟ ਅਲ ਦੁਆਰਾ ਵਰਣਿਤ ਵਿਧੀ ਦੀ ਵਰਤੋਂ ਕੀਤੀ।34. ਸੰਖੇਪ ਵਿੱਚ, ਲਿਆਂਗ ਐਟ ਅਲ ਦੁਆਰਾ ਦੱਸੇ ਗਏ ਦਿਸ਼ਾ-ਨਿਰਦੇਸ਼ਾਂ ਅਨੁਸਾਰ ਚਾਰ sgRNAs ਦੀ ਚੋਣ ਕੀਤੀ ਗਈ ਸੀ।35. CC-Top CRISPR/Cas9 ਟਾਰਗੇਟ ਪੂਰਵ ਅਨੁਮਾਨ ਟੂਲ ਦੀ ਵਰਤੋਂ sgRNAs ਦੀ ਸੂਚੀ ਬਣਾਉਣ ਲਈ ਕੀਤੀ ਗਈ ਸੀ36, Mfold ਵੈੱਬ ਸਰਵਰ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਫੋਲਡਿੰਗ ਦਾ ਮੁਲਾਂਕਣ ਕੀਤਾ ਗਿਆ ਸੀ37, ਅਤੇ sgRNAs ਦੀ ਗਤੀਵਿਧੀ ਦੀ sgRNA ਸਕੋਰਰ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਭਵਿੱਖਬਾਣੀ ਕੀਤੀ ਗਈ ਸੀ38. ਨਿਮਨਲਿਖਤ ਗਾਈਡਾਂ ਨੂੰ ਚੁਣਿਆ ਗਿਆ ਸੀ ਅਤੇ ਵੈਕਟਰ pAGM:CRISPRASli: exon5.1T01 (ATTTCATCCGCGATCTCTCGGGG), exon5.1T04 (GATTTCA TCCGCGATCTCTCGGG), exon5.1T06 (TATTTCCTATTGCTACCAGATTGTCG5.1) ਅਤੇ exon07T86966 (TATTTCCTATTTGCTACCAGATTGTCG01009), ਅਤੇ। CRISPR ਕੰਸਟਰੱਕਟ ਨੂੰ ਫਿਰ Addgene: pICH0 (ਐਂਪਲੀਫਿਕੇਸ਼ਨ ਲਈ ਟੈਪਲੇਟ) ਤੋਂ ਪ੍ਰਾਪਤ ਪਲਾਜ਼ਮੀਡਾਂ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਸੰਸਲੇਸ਼ਣ ਕੀਤਾ ਗਿਆ ਸੀ; pICSL47751 (ਪੱਧਰ 47761 ਪਲਾਜ਼ਮੀਡ); pICH47772, pICH47781, pICH47732, pICH1 ਅਤੇ pICH41822 (ਪੱਧਰ 4723 ਪਲਾਜ਼ਮੀਡ); piCH2 (ਚਾਰ ਗਾਈਡਾਂ ਲਈ ਲਿੰਕਰ); ਅਤੇ pAGMXNUMX (ਪੱਧਰ XNUMX ਬਾਈਨਰੀ ਵੈਕਟਰ)। ਪਲਾਜ਼ਮੀਡ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਕਲੋਨ ਕੀਤਾ ਗਿਆ ਸੀ ਈ. ਕੋਲਾਈ DH5a ਅਤੇ ਸ਼ੁੱਧ ਪਲਾਜ਼ਮਿਡ ਨੂੰ ਪ੍ਰਾਈਮਰ PDS5843 (TTTGTGATGCTCGTCGGGGG), PDS8535 (CCCGAGAATTTATGCAGCATT TT) PDS8536 (TCATCAGTCAATTATGCAGCATT TT) PDS717 (TCATCAGTCAATTATTACGGGGGCT), ਅਤੇ AL9 (GCCTTCAGXNUMX) ਦੀ ਮੌਜੂਦਗੀ ਦੀ ਪੁਸ਼ਟੀ ਕਰਨ ਲਈ ਪ੍ਰਾਈਮਰਾਂ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਕ੍ਰਮਬੱਧ ਕਰਨ ਲਈ ਭੇਜਿਆ ਗਿਆ ਸੀ।
pBINPLUS-Sli ਅਤੇ pAGMzCRISPR ਦਾ ਪਰਿਵਰਤਨAਵੈਕਟਰ ਵਿੱਚ ਸਲਾਈ ਕਰੋ ਐਗਰੋਬੈਕੈਕਟੀਰੀਅਮ. ਅਸੀਂ pBINPLUS-Sli ਨੂੰ ਵਿੱਚ ਬਦਲ ਦਿੱਤਾ ਹੈ A. tumefaciens AGL0 ਅਤੇ pAGM:CRISPRASli ਨੂੰ A ਵਿੱਚ ਦਬਾਓ। tumefaciens ਇੱਕ ਇਲੈਕਟ੍ਰੋਪੋਰੇਸ਼ਨ ਪ੍ਰੋਟੋਕੋਲ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ AGL0 ਅਤੇ AGL1 ਨੂੰ ਤਣਾਅ ਕਰਦਾ ਹੈ। ਅਸੀਂ 40 PL ਸਮਰੱਥ AGL0 ਸੈੱਲ ਲਏ ਅਤੇ 110 pl ਬਰਫ਼-ਠੰਡੇ milliQ ਪਾਣੀ ਨੂੰ ਜੋੜਿਆ। ਅਸੀਂ ਇਸ ਮਿਸ਼ਰਣ ਦੇ 50 pl ਨੂੰ ਬਰਫ਼ 'ਤੇ ਪ੍ਰੀ-ਕੂਲਡ ਐਪੇਨਡੋਰਫ ਟਿਊਬਾਂ ਵਿੱਚ ਪਾਈਪ ਕੀਤਾ ਅਤੇ ਪਲਾਜ਼ਮੀਡ ਦਾ 1 pl ਜੋੜਿਆ। ਅਸੀਂ ਸੈੱਲਾਂ ਨੂੰ 15 ਮਿੰਟ ਲਈ ਬਰਫ਼ 'ਤੇ ਛੱਡ ਦਿੱਤਾ ਅਤੇ ਸੈੱਲਾਂ ਨੂੰ ਪ੍ਰੀ-ਕੂਲਡ ਇਲੈਕਟ੍ਰੋਪੋਰੇਸ਼ਨ ਕੁਵੇਟਸ ਵਿੱਚ ਤਬਦੀਲ ਕਰ ਦਿੱਤਾ। ਅਸੀਂ Ec1 (1.8 kV, 0.1 ਸੈਂਟੀਮੀਟਰ ਕਯੂਵੇਟ) ਪ੍ਰੋਗਰਾਮ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਮਿਸ਼ਰਣਾਂ ਨੂੰ ਮਾਈਕ੍ਰੋਪੁਲਸਰ™ (ਬਾਇਓ-ਰੈਡ ਲੈਬਾਰਟਰੀਜ਼, ਵੀ-ਨੇਂਡਾਲ, ਨੀਦਰਲੈਂਡਜ਼) ਨਾਲ ਇਲੈਕਟ੍ਰੋਪੋਰੇਟ ਕੀਤਾ। ਅਸੀਂ 1 ਮਿਲੀਲੀਟਰ LB ਜੋੜਿਆ ਅਤੇ ਸੈੱਲਾਂ ਨੂੰ 3 ° C ਅਤੇ 28 RPM 'ਤੇ ਸ਼ੇਕਰ 'ਤੇ 200 ਘੰਟੇ ਲਈ ਪ੍ਰਫੁੱਲਤ ਕੀਤਾ। ਬਾਅਦ ਵਿੱਚ, ਅਸੀਂ ਪਰਿਵਰਤਨ ਸੰਸਕ੍ਰਿਤੀ ਦੇ ਨਾਲ ਰਿਫੈਮਪਿਸਿਨ (100 pg/ml) ਅਤੇ Kanamycin (50 pg/ml) ਵਾਲੀਆਂ LB ਅਗਰ ਪਲੇਟਾਂ ਦਾ ਟੀਕਾ ਲਗਾਇਆ। ਸਾਰੀਆਂ ਚੁਣੀਆਂ ਗਈਆਂ ਕਲੋਨੀਆਂ ਨੂੰ ਸਹੀ ਵੈਕਟਰ ਰੱਖਣ ਦੀ ਪੁਸ਼ਟੀ ਕੀਤੀ ਗਈ ਸੀ।
ਆਲੂ ਜੀਨੋਟਾਈਪ ਦਾ ਪਰਿਵਰਤਨ. ਅਸੀਂ ਜੀਨੋਟਾਈਪ B666 ਅਤੇ B667 ਨੂੰ pBINPLUS-Sli ਨਾਲ ਅਤੇ ਜੀਨੋਟਾਈਪ B663 ਅਤੇ B665 ਨੂੰ pAGM:CRISPRASli ਵੈਕਟਰ ਨਾਲ ਵਿਸਰ ਦੁਆਰਾ ਵਰਣਿਤ ਸਟੈਮ ਐਕਸਪਲਾਂਟ ਵਿਧੀ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਬਦਲਿਆ ਹੈ।39. ਸੰਖੇਪ ਰੂਪ ਵਿੱਚ, ਇੰਟਰਨੋਡ ਐਕਸਪਲਾਂਟ ਇਨ ਵਿਟਰੋ ਵਧੇ ਹੋਏ ਜੀਨੋਟਾਈਪਾਂ ਤੋਂ ਪ੍ਰਾਪਤ ਕੀਤੇ ਗਏ ਸਨ ਅਤੇ PACM ਮਾਧਿਅਮ ਦੇ 3 ਮਿ.ਲੀ. ਦੇ ਨਾਲ R2B ਮਾਧਿਅਮ ਵਾਲੇ ਪੈਟਰੀ ਪਕਵਾਨਾਂ 'ਤੇ ਪਾਏ ਗਏ ਸਨ। ਅਗਲੇ ਦਿਨ, 50 ਐੱਚ. ਦੇ 48 ਮਿ.ਲੀ ਐਗਰੋਬੈਕਟੀਰੀਅਮ ਸਭਿਆਚਾਰਾਂ ਨੂੰ ਕੇਂਦਰਿਤ ਕੀਤਾ ਗਿਆ ਸੀ ਅਤੇ LB ਦੇ 75 ਮਿਲੀਲੀਟਰ ਵਿੱਚ ਮੁੜ ਮੁਅੱਤਲ ਕੀਤਾ ਗਿਆ ਸੀ। ਇੰਟਰਨੋਡ ਐਕਸਪਲਾਂਟ ਫਿਰ ਵਿੱਚ ਡੁੱਬ ਗਏ ਸਨ ਐਗਰੋਬੈਕਟੀਰੀਅਮ 5 ਮਿੰਟ ਲਈ ਮੁਅੱਤਲ, ਫਿਲਟਰ 'ਤੇ ਸੁੱਕਿਆ, ਅਤੇ R3B ਮੀਡੀਅਮ ਵਾਲੇ ਪੈਟਰੀ ਪਕਵਾਨਾਂ 'ਤੇ ਵਾਪਸ ਰੱਖਿਆ ਗਿਆ। 48 ਘੰਟਿਆਂ ਦੇ ਪ੍ਰਫੁੱਲਤ ਹੋਣ ਤੋਂ ਬਾਅਦ, ਐਕਸਪਲਾਂਟ ਨੂੰ ਐਂਟੀਬਾਇਓਟਿਕਸ ਦੇ ਨਾਲ MS20 ਵਾਲੇ ਪੈਟਰੀ ਪਕਵਾਨਾਂ ਵਿੱਚ ਤਬਦੀਲ ਕਰ ਦਿੱਤਾ ਗਿਆ ਅਤੇ ਕਮਤ ਵਧਣੀ ਦੇ ਪੁਨਰਜਨਮ ਦੀ ਆਗਿਆ ਦੇਣ ਲਈ ਇੱਕ ਵਿਕਾਸ ਚੈਂਬਰ ਵਿੱਚ ਰੱਖਿਆ ਗਿਆ। ਪੁਨਰਜਨਮ ਤੋਂ ਬਾਅਦ, ਕਮਤ ਵਧਣੀ MS20 ਮੀਡੀਆ 'ਤੇ ਉਗਾਈ ਗਈ ਸੀ ਜਿਸ ਵਿੱਚ ਸੇਫੋਟੈਕਸਾਈਮ (200 pg/ml), ਵੈਨਕੋਮਾਈਸਿਨ (200 pg/ml), ਅਤੇ kanamycin (100 pg/ml) ਸ਼ਾਮਲ ਸਨ। ਜਦੋਂ ਕਮਤ ਵਧਣੀ ਲੋੜੀਂਦੀ ਲੰਬਾਈ 'ਤੇ ਪਹੁੰਚ ਗਈ, ਤਾਂ ਕਟਿੰਗਜ਼ ਬਣਾਈਆਂ ਗਈਆਂ ਅਤੇ ਬਿਨਾਂ ਐਂਟੀਬਾਇਓਟਿਕਸ ਦੇ MS20 ਵਿੱਚ ਉਗਾਈਆਂ ਗਈਆਂ। ਘੱਟੋ-ਘੱਟ ਦੋ ਹਫ਼ਤਿਆਂ ਬਾਅਦ ਐਂਟੀਬਾਇਓਟਿਕਸ ਤੋਂ ਬਿਨਾਂ MS20 ਦੇ ਵਧਣ ਤੋਂ ਬਾਅਦ, ਪੌਦੇ ਗ੍ਰੀਨਹਾਉਸ ਵਿੱਚ ਲਗਾਏ ਗਏ ਸਨ।
ਪਲੋਡੀ ਵਿਸ਼ਲੇਸ਼ਣ. ਪਲਾਂਟ ਸਾਇਟੋਮੈਟਰੀ ਸਰਵਿਸਿਜ਼ (ਡਿਦਾਮ, ਨੀਦਰਲੈਂਡਜ਼) ਦੁਆਰਾ ਪ੍ਰਵਾਹ ਸਾਇਟੋਮੈਟਰੀ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਟ੍ਰਾਂਸਜੇਨਿਕ ਪੌਦਿਆਂ ਦੇ ਨਾਲ-ਨਾਲ ਗੈਰ-ਪਰਿਵਰਤਿਤ ਨਿਯੰਤਰਣਾਂ ਦੀ ਚਾਲ ਨਿਰਧਾਰਤ ਕੀਤੀ ਗਈ ਸੀ। ਸਾਰੇ ਟੈਟਰਾਪਲੋਇਡ ਰੀਜਨਰੈਂਟਸ ਨੂੰ ਰੱਦ ਕਰ ਦਿੱਤਾ ਗਿਆ ਸੀ।
CRISPR ਦਾ ਪੇਜ ਵਿਸ਼ਲੇਸ਼ਣ-Cas9-ਪ੍ਰੇਰਿਤ ਪਰਿਵਰਤਨ. ਡੀਐਨਏ ਕੱਢਣ, ਪੀਸੀਆਰ, ਅਤੇ ਪੇਜ ਵਿਸ਼ਲੇਸ਼ਣ ਲਿਮਗਰੁੱਪ (ਹੋਰਸਟ, ਨੀਦਰਲੈਂਡਜ਼) ਦੁਆਰਾ ਕੀਤੇ ਗਏ ਸਨ। ਹੇਠਾਂ ਦਿੱਤੇ ਪ੍ਰਾਈਮਰਾਂ ਦੀ ਵਰਤੋਂ CRISPR-Cas9-ਨਿਸ਼ਾਨਾ ਖੇਤਰ ਨੂੰ ਵਧਾਉਣ ਲਈ ਕੀਤੀ ਗਈ ਸੀ: ਫਾਰਵਰਡ ਪ੍ਰਾਈਮਰ: CTATTTCCTATTGCTACCAG, ਰਿਵਰਸ ਪ੍ਰਾਈਮਰ: AAACTTTACCCAAAT AACGTC। PCR ਉਤਪਾਦਾਂ ਦੀ ਲੇਬਲਿੰਗ ਇੱਕ M13 ਟੇਲ (ਪ੍ਰਾਈਮਰ ਕ੍ਰਮ: TGTAAAACGACGCAGTAAACTTTAC CCAAATAACGTC) ਅਤੇ PCR ਮਿਸ਼ਰਣ ਵਿੱਚ 700 IRDye ਜਾਂ 800 IRDye ਲੇਬਲ ਵਾਲੇ M13 ਪ੍ਰਾਈਮਰ ਦੇ ਨਾਲ ਇੱਕ ਉਲਟਾ ਪ੍ਰਾਈਮਰ ਜੋੜ ਕੇ ਪ੍ਰਾਪਤ ਕੀਤੀ ਗਈ ਸੀ। ਨਤੀਜੇ ਵਜੋਂ PCR ਉਤਪਾਦਾਂ ਦਾ ਇੱਕ Li-cor ਸਿਸਟਮ ਦੀ ਵਰਤੋਂ ਕਰਕੇ PAGE 'ਤੇ ਵਿਸ਼ਲੇਸ਼ਣ ਕੀਤਾ ਗਿਆ ਸੀ। CRISPR-Cas9-ਪ੍ਰੇਰਿਤ ਪਰਿਵਰਤਨ ਤੋਂ ਬਿਨਾਂ ਜ਼ਿਆਦਾਤਰ ਲਾਈਨਾਂ ਦੇ ਪੀਸੀਆਰ ਉਤਪਾਦਾਂ ਨੂੰ ਚਿੱਤਰ ਬਣਾਉਣ ਲਈ ਜੈੱਲ ਚਿੱਤਰਾਂ ਤੋਂ ਬਾਹਰ ਕੱਢਿਆ ਗਿਆ ਸੀ। 3d (ਡੈਸ਼ਡ ਲਾਈਨਾਂ ਦੁਆਰਾ ਦਰਸਾਏ ਐਕਸਾਈਜ਼)।
533 ਬੀਪੀ ਸੰਮਿਲਨ ਦਾ ਫਾਈਲੋਜੈਨੇਟਿਕ ਵਿਸ਼ਲੇਸ਼ਣ. NCBI ਵੈੱਬਸਾਈਟ 'ਤੇ BLASTn ਦੀ ਵਰਤੋਂ ਕਰਕੇ 533 bp ਸੰਮਿਲਨ ਦੇ ਕ੍ਰਮ ਦਾ ਵਿਸ਼ਲੇਸ਼ਣ ਕੀਤਾ ਗਿਆ ਸੀ। ਵਿੱਚ ਮੌਜੂਦ ਸੰਮਿਲਨ ਸਮੇਤ ਸਭ ਤੋਂ ਵਧੀਆ 30 ਹਿੱਟ ਸਲੀ Solyntus ਵਿੱਚ, ਫਿਰ ਡਿਫੌਲਟ ਸੈਟਿੰਗਾਂ ਦੇ ਨਾਲ MUSCLE ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ MegAlign Pro 17 (DNASTAR) ਵਿੱਚ ਡਾਊਨਲੋਡ ਅਤੇ ਅਲਾਈਨ ਕੀਤੇ ਗਏ ਸਨ। ਰੁੱਖਾਂ ਨੂੰ ਡਿਫੌਲਟ ਸੈਟਿੰਗਾਂ ਦੇ ਨਾਲ ਗੁਆਂਢੀ-ਜੋੜਨ ਵਾਲੇ ਐਲਗੋਰਿਦਮ ਦੀ ਵਰਤੋਂ ਕਰਕੇ ਤਿਆਰ ਕੀਤਾ ਗਿਆ ਸੀ।
ਰਿਪੋਰਟਿੰਗ ਸੰਖੇਪ. ਖੋਜ ਡਿਜ਼ਾਈਨ ਬਾਰੇ ਹੋਰ ਜਾਣਕਾਰੀ ਇਸ ਲੇਖ ਨਾਲ ਜੁੜੇ ਕੁਦਰਤ ਖੋਜ ਰਿਪੋਰਟਿੰਗ ਸੰਖੇਪ ਵਿੱਚ ਉਪਲਬਧ ਹੈ।
ਡਾਟਾ ਉਪਲਬਧਤਾ
ਸੋਲਿੰਟਸ ਜੀਨੋਮ ਕ੍ਰਮ ਅਤੇ ਕੱਚੇ ਕ੍ਰਮ ਰੀਡਜ਼ NCBI 'ਤੇ ਪਹੁੰਚ ਅਧੀਨ ਉਪਲਬਧ ਹਨ PRJNA631911. ਸੋਲਿੰਟਸ ਜੀਨੋਮ ਅਸੈਂਬਲੀ ਅਤੇ ਐਨੋਟੇਸ਼ਨ ਫਾਈਲਾਂ WUR ਤੋਂ ਉਪਲਬਧ ਹਨ [https://www.plantbreeding.wur.nl/Solyntus/]. ਉਗਣ ਵਾਲੇ ਪਰਾਗ ਤੋਂ ਆਰਐਨਏ ਸੀਕੁਏਂਸਿੰਗ ਡੇਟਾ NCBI ਸ਼ਾਰਟ ਰੀਡ ਆਰਕਾਈਵ ਅੰਡਰ ਐਕਸੈਸ਼ਨ 'ਤੇ ਉਪਲਬਧ ਹੈ। PRJNA713577. ਹੋਰ ਡੇਟਾ ਸਰੋਤ ਡੇਟਾ ਫਾਈਲ ਵਿੱਚ ਉਪਲਬਧ ਹੈ ਜਾਂ ਬੇਨਤੀ ਕਰਨ 'ਤੇ ਉਪਲਬਧ ਕਰਾਇਆ ਜਾਵੇਗਾ। ਸਰੋਤ ਡੇਟਾ ਇਸ ਪੇਪਰ ਦੇ ਨਾਲ ਪ੍ਰਦਾਨ ਕੀਤੇ ਗਏ ਹਨ.
ਪ੍ਰਾਪਤ ਹੋਇਆ: 22 ਜਨਵਰੀ 2021; ਸਵੀਕਾਰ ਕੀਤਾ: 8 ਜੂਨ 2021;
ਆਨਲਾਈਨ ਪ੍ਰਕਾਸ਼ਿਤ: 06 ਜੁਲਾਈ 2021
ਹਵਾਲੇ
- 1. ਡੁਵਿਕ, DN ਮੱਕੀ (ਜ਼ੀਆ) ਵਿੱਚ ਉੱਨਤੀ ਪੈਦਾ ਕਰਨ ਲਈ ਪ੍ਰਜਨਨ ਦਾ ਯੋਗਦਾਨ ਮਈ ਐੱਲ.). ਐਡਵੋਕੇਟ ਐਗਰੋਨ। 86, 83-145 (2005)।
- 2. Lindhout, P. et al. F1 ਹਾਈਬ੍ਰਿਡ ਬੀਜ ਆਲੂ ਦੇ ਪ੍ਰਜਨਨ ਵੱਲ। ਆਲੂ Res. 54, 301-312 (2011)।
- 3. ਜੈਨਸਕੀ, ਐਸਐਚ ਐਟ ਅਲ. ਆਲੂ ਨੂੰ ਇੱਕ ਡਿਪਲੋਇਡ ਇਨਬ੍ਰੇਡ ਲਾਈਨ-ਆਧਾਰਿਤ ਫਸਲ ਵਜੋਂ ਮੁੜ ਖੋਜਣਾ। ਫਸਲ ਵਿਗਿਆਨ. 56, 1412-1422 (2016)।
- 4. ਯੇ, ਐੱਮ. ਐਟ ਅਲ. S-RNase ਦੇ ਨਾਕਆਊਟ ਦੁਆਰਾ ਸਵੈ-ਅਨੁਕੂਲ ਡਿਪਲੋਇਡ ਆਲੂ ਦਾ ਉਤਪਾਦਨ। ਨੈਟ. ਪੌਦੇ 4, 651-654 (2018)।
- 5. Enciso-Rodriguez, F. et al. CRISPR-cas9 ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਡਿਪਲੋਇਡ ਆਲੂ ਵਿੱਚ ਸਵੈ-ਅਸੰਗਤਤਾ ਨੂੰ ਦੂਰ ਕਰਨਾ। ਸਾਹਮਣੇ ਪੌਦਾ ਵਿਗਿਆਨ. 10, 1-12 (2019)।
- 6. Su, Y. et al. ਦੇ ਵਿਰੁੱਧ ਪ੍ਰਤੀਰੋਧ ਲਈ ਜੀਨਾਂ ਦੀ ਘੁਸਪੈਠ ਫਾਈਟਰਥੋਥਰਾ ਇਨਫੈਸਟਨ ਡਿਪਲੋਇਡ ਆਲੂ ਵਿੱਚ. ਐਮ. ਜੇ. ਆਲੂ ਰੈਸ. 97, 33-42 (2020)।
- 7. ਡਿਪਲੋਇਡ ਆਲੂ ਸਪੀਸੀਜ਼ ਵਿੱਚ ਡੀਜ਼ਿਡਜ਼ੀਨਿਓ, ਡੀਕੇ, ਬ੍ਰਾਇਨ, ਜੀਜੇ, ਵਾਈਲਡ, ਜੀ. ਅਤੇ ਰੌਬਿਨਸ, ਟੀਪੀ ਐਲੇਲਿਕ ਵਿਭਿੰਨਤਾ ਐਸ-ਆਰਨੇਜ਼ ਐਲੀਲਜ਼ ਦੀ। ਥਿਊਰ. ਐਪਲ. ਜੈਨੇਟ. 129, 1985-2001 (2016)।
- 8. McClure, B., Cruz-Garcia, F. & Romero, C. S-RNase-ਅਧਾਰਿਤ ਪ੍ਰਣਾਲੀਆਂ ਵਿੱਚ ਅਨੁਕੂਲਤਾ ਅਤੇ ਅਸੰਗਤਤਾ। ਐਨ. ਬੋਟ. 108, 647-658 (2011)।
- 9. ਕੁਬੋ, ਕੇ. ਐਟ ਅਲ. S-RNase-ਅਧਾਰਿਤ ਸਵੈ-ਅਸੰਗਤਤਾ ਵਿੱਚ ਸਹਿਯੋਗੀ ਗੈਰ-ਸਵੈ-ਪਛਾਣ ਪ੍ਰਣਾਲੀ। ਸਾਇੰਸ 330, 796-799 (2010)।
- 10. ਡੀ ਜੋਂਗ, ਐਚ. ਅਤੇ ਰੋਵੇ, ਕਾਸ਼ਤ ਕੀਤੇ ਡਿਪਲੋਇਡ ਆਲੂਆਂ ਵਿੱਚ ਪੀਆਰ ਇਨਬ੍ਰੀਡਿੰਗ। ਆਲੂ Res. 14, 74-83 (1971)।
- 11. ਹਰਮਸਨ, ਜੇ.ਜੀ.ਟੀ. ਅਤੇ ਓਲਡਰ, ਜੇ. ਡਾਇਹਾਪਲੋਇਡਜ਼ ਵਿੱਚ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਦੇ ਜੈਨੇਟਿਕਸ ਸੋਲਨਮ ਟਿosਬਰੋਸਮ L. 1. ਦੋ ਸਵੈ-ਅਨੁਕੂਲ ਡਾਇਹਾਪਲੋਇਡਜ਼ ਦਾ ਪ੍ਰਜਨਨ ਵਿਵਹਾਰ। ਯੂਫਾਈਟਿਕਾ 25, 597-607 (1976)।
- 12. ਹੋਸਾਕਾ, ਕੇ. ਅਤੇ ਹੈਨੇਮਨ, ਆਰ.ਈ. ਜੂਨੀਅਰ ਸਵੈ-ਅਨੁਕੂਲ ਜੰਗਲੀ ਡਿਪਲੋਇਡ ਆਲੂ ਸਪੀਸੀਜ਼ ਵਿੱਚ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਦੇ ਜੈਨੇਟਿਕਸ ਸੋਲਨਮ ਚੈਕੋਏਂਸ. 1. ਇੱਕ S ਲੋਕਸ ਇਨਿਹਿਬਟਰ (Sli) ਜੀਨ ਦਾ ਪਤਾ ਲਗਾਉਣਾ। ਯੂਫਾਈਟਿਕਾ 99, 191-197 (1998)।
- 13. ਹੋਸਾਕਾ, ਕੇ. ਅਤੇ ਹੈਨੇਮਨ, ਆਰ.ਈ. ਜੂਨੀਅਰ ਸਵੈ-ਅਨੁਕੂਲ ਜੰਗਲੀ ਡਿਪਲੋਇਡ ਆਲੂ ਸਪੀਸੀਜ਼ ਵਿੱਚ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਦੇ ਜੈਨੇਟਿਕਸ ਸੋਲਨਮ ਚੈਕੋਏਂਸ. 2. ਡੀਐਨਏ ਮਾਰਕਰ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਆਲੂ ਦੇ ਜੀਨੋਮ 'ਤੇ ਇੱਕ ਐਸ ਲੋਕਸ ਇਨਿਹਿਬਟਰ (Sli) ਜੀਨ ਦਾ ਸਥਾਨੀਕਰਨ। ਯੂਫਾਈਟਿਕਾ 103, 265-271 (1998)।
- 14. ਬਿਰਹਮਨ, ਆਰ.ਕੇ. ਅਤੇ ਹੋਸਾਕਾ, ਕੇ. ਇੱਕ ਐਸ-ਲੋਕਸ ਇਨਿਹਿਬਟਰ (ਸਲੀ) ਜੀਨ ਦੀ ਵਰਤੋਂ ਕਰਦੇ ਹੋਏ ਡਿਪਲੋਇਡ ਆਲੂਆਂ ਦੀਆਂ ਨਸਲਾਂ ਦਾ ਉਤਪਾਦਨ, ਅਤੇ ਉਹਨਾਂ ਦੀ ਵਿਸ਼ੇਸ਼ਤਾ। ਜੀਨੋਮ 502, 495-502 (2000)।
- 15. ਚੇਨ, ਜੇ., ਹੂ, ਕਿਊ., ਝਾਂਗ, ਵਾਈ., ਲੂ, ਸੀ. ਅਤੇ ਕੁਆਂਗ, ਐਚ. ਪੀ-ਮਾਈਟ: ਪੌਦਿਆਂ ਦੇ ਲਘੂ ਇਨਵਰਟੇਡ-ਰਿਪੀਟ ਟ੍ਰਾਂਸਪੋਸੇਬਲ ਤੱਤਾਂ ਲਈ ਇੱਕ ਡੇਟਾਬੇਸ। ਨਿਊਕਲੀਐਸਿਜ਼ ਐੱਸ. 42, 1176-1181 (2014)।
- 16. ਸਟੇਫਾਨੋਵਿਕਜ਼, ਕੇ., ਲੈਨਨੂ, ਐਨ. ਅਤੇ ਵੈਨ ਡੈਮੇ, ਈਜੇਐਮ ਪਲਾਂਟ ਐਫ-ਬਾਕਸ ਪ੍ਰੋਟੀਨ- ਜੀਵਨ ਅਤੇ ਮੌਤ ਵਿਚਕਾਰ ਨਿਰਣਾਇਕ। ਕ੍ਰਿਟ. ਰੇਵ. ਪਲਾਂਟ ਸਾਇੰਸ. 34, 523-552 (2015)।
- 17. ਲਿਊ, ਬੀ., ਮੋਰਸ, ਡੀ. ਅਤੇ ਕੈਪਾਡੋਸੀਆ, ਐੱਮ. S-RNases ਦਾ ਗਲਾਈਕੋਸੀਲੇਸ਼ਨ ਸੋਲਨਮ ਚੈਕੋਏਂਸ ਵਿੱਚ ਪਰਾਗ ਰੱਦ ਕਰਨ ਦੇ ਥ੍ਰੈਸ਼ਹੋਲਡ ਨੂੰ ਪ੍ਰਭਾਵਿਤ ਕਰ ਸਕਦਾ ਹੈ। ਜੇ. ਐਕਸਪ. ਬੋਟ. 59, 545-552 (2008)।
- 18. ਲੇਮਬੀਰ, ਐਫਪੀਈ ਆਲੂ ਜੀਨੋਮਿਕਸ ਤਿੰਨ ਤਰੀਕੇ: ਕੰਦਾਂ ਵਿੱਚ ਐਂਡੋਰਡਡੁਪਲੀਕੇਸ਼ਨ ਦੀ ਮਾਤਰਾ, ਟ੍ਰਾਂਸਪੋਸਨ ਭੂਮੀ ਦੁਆਰਾ ਇੱਕ ਰੋੰਪ, ਅਤੇ ਫੁੱਲਾਂ ਦੇ ਰੰਗ ਨਿਯਮ ਦੀ ਵਿਆਖਿਆ। http://hdl.handle.net/10919/84480 (2018).
- 19. ਕਲਾਟ, ਸੀਆਰ ਐਟ ਅਲ. ਆਲੂ ਵਿੱਚ ਸਲੀ-ਅਧਾਰਿਤ ਸਵੈ-ਅਨੁਕੂਲਤਾ ਦੀ ਉਤਪੱਤੀ ਅਤੇ ਵਿਆਪਕ ਮੌਜੂਦਗੀ। ਥਿਊਰ. ਐਪਲ. ਜੈਨੇਟ. https://doi.org/10.1007/s00122-020-03627-8 (2020).
- 20. ਐਂਡਲਮੈਨ, ਜੇ., ਜੈਨਸਕੀ, ਐਸ.ਐਚ., ਬਟਲਰ, ਐਨ. ਅਤੇ ਕ੍ਰਿਸਟੇਨਸਨ, ਜੀ. ਆਲੂ ਦੇ ਕ੍ਰੋਮੋਸੋਮ 'ਤੇ ਇੱਕ ਵਿਗਾੜ ਵਾਲੇ ਘਾਤਕ ਐਲੀਲ ਦੇ ਜੈਨੇਟਿਕ ਸਬੂਤ ਅਮਰੀਕਾ ਦੇ ਪੋਟੇਟੋ ਐਸੋਸੀਏਸ਼ਨ ਦੀ 12-ਸਲਾਨਾ ਰਿਪੋਰਟ। ਐਮ. ਜੇ. ਆਲੂ ਰੈਸ. 96, 331 (2019).
- 21. ਵੈਨ ਲਿਸ਼ੌਟ, ਐਨ. ਐਟ ਅਲ. ਸੋਲਿੰਟਸ, ਆਲੂ (ਸੋਲਨਮ ਟਿਊਬਰੋਜ਼ਮ) ਲਈ ਨਵਾਂ ਬਹੁਤ ਹੀ ਅਨੁਕੂਲ ਹਵਾਲਾ ਜੀਨੋਮ। G3 ਜੀਨਸ ਜੀਨੋਮ ਜੈਨੇਟ. 10, 3489-3495 (2020)।
- 22. Leisner, CP et al. M6 ਦਾ ਜੀਨੋਮ ਕ੍ਰਮ, ਉੱਚ-ਗਲਾਈਕੋਆਕਲਾਇਡ-ਉਤਪਾਦਕ ਕੰਦ-ਬੇਅਰਿੰਗ ਆਲੂ ਸਪੀਸੀਜ਼ ਦਾ ਇੱਕ ਡਿਪਲੋਇਡ ਇਨਬ੍ਰੇਡ ਕਲੋਨ ਸੋਲਨਮ ਚੈਕੋਏਂਸ, ਬਕਾਇਆ ਹੈਟਰੋਜ਼ਾਈਗੋਸਿਟੀ ਨੂੰ ਪ੍ਰਗਟ ਕਰਦਾ ਹੈ। ਪਲਾਂਟ ਜੇ. 1967, 562-570 (2018)।
- 23. ਪੀਟਰਸਨ, ਬੀਏ ਐਟ ਅਲ. ਇਨਫਿਨਿਅਮ 8303 ਆਲੂ ਸਿੰਗਲ-ਨਿਊਕਲੀਓਟਾਈਡ ਪੋਲੀਮੋਰਫਿਜ਼ਮ ਐਰੇ ਨਾਲ ਪਰੀਖਣ ਕੀਤੇ ਗਏ ਇੱਕ ਕਾਸ਼ਤ ਕੀਤੇ ਡਿਪਲੋਇਡ ਆਲੂ ਦੀ ਆਬਾਦੀ ਵਿੱਚ ਸਵੈ-ਉਪਜਾਊ ਸ਼ਕਤੀ। ਪੌਦਾ ਜੀਨੋਮ https://doi.org/10.3835/plantgenome2016.01.0003 (2016).
- 24. ਲਿਆਨ, ਕਿਊ. ਐਟ ਅਲ. ਆਲੂ ਪੌਲੀਪਲੋਇਡਾਈਜ਼ੇਸ਼ਨ ਦੇ ਦੌਰਾਨ ਨੁਕਸਾਨਦੇਹ ਪਰਿਵਰਤਨ ਦੀ ਪ੍ਰਾਪਤੀ. ਜੇ. ਇੰਟੀਗਰ ਪੌਦਾ ਬਾਇਓਲ. 61, 7-11 (2019)।
- 25. ਵੈਨ ਲਿਸ਼ਆਉਟ, ਐਨ. ਐਟ ਅਲ. ਸੋਲਿੰਟਸ, ਆਲੂ ਲਈ ਨਵਾਂ ਬਹੁਤ ਹੀ ਅਨੁਕੂਲ ਹਵਾਲਾ ਜੀਨੋਮ (ਸੋਲੇਨਮ ਟਿਊਬਰੋਸਮ)। G3 ਜੀਨਸ ਜੀਨੋਮ ਜੈਨੇਟ. 631911, g3.401550.2020 (2020)।
- 26. ਵੈਨ ਓਈਜੇਨ, ਜੇਡਬਲਯੂ JoinMap®4, ਪ੍ਰਯੋਗਾਤਮਕ ਆਬਾਦੀ ਵਿੱਚ ਜੈਨੇਟਿਕ ਲਿੰਕੇਜ ਨਕਸ਼ਿਆਂ ਦੀ ਗਣਨਾ ਲਈ ਸੌਫਟਵੇਅਰ ਵੋਲ. 33 (ਕਿਆਜ਼ਮਾ ਬੀਵੀ, 2006)।
- 27. ਵੈਨ ਓਈਜੇਨ, ਆਟੋਗੈਮਸ ਸਪੀਸੀਜ਼ ਵਿੱਚ ਮਾਤਰਾਤਮਕ ਗੁਣ ਸਥਾਨ ਦੀ ਮੈਪਿੰਗ ਦੀ JW ਸ਼ੁੱਧਤਾ। ਥਿਊਰ. ਐਪਲ ਜੈਨੇਟ. 84, 803-811 (1992)।
- 28. ਵੂਰਿਪਸ, RE ਮੈਪਚਾਰਟ: ਲਿੰਕੇਜ ਨਕਸ਼ਿਆਂ ਅਤੇ QTLs ਦੀ ਗ੍ਰਾਫਿਕਲ ਪੇਸ਼ਕਾਰੀ ਲਈ ਸੌਫਟਵੇਅਰ। ਜੇ. ਹੇਰਡ 93, 77-78 (2002)।
- 29. Hirsch, CD et al. ਸਪੂਡ ਡੀਬੀ: ਆਲੂ ਦੇ ਪ੍ਰਜਨਨ ਨੂੰ ਤੇਜ਼ ਕਰਨ ਲਈ ਮਾਈਨਿੰਗ ਕ੍ਰਮ, ਜੀਨੋਟਾਈਪ ਅਤੇ ਫੀਨੋਟਾਈਪ ਲਈ ਇੱਕ ਸਰੋਤ। ਪੌਦਾ ਜੀਨੋਮ. 7, https://doi.org/ 10.3835/ਪਲਾਂਟਜੀਨੋਮ2013.12.0042 (2014).
- 30. ਬੈਂਕੇਵਿਚ, ਏ. ਏਟ ਅਲ. SPAdes: ਇੱਕ ਨਵਾਂ ਜੀਨੋਮ ਅਸੈਂਬਲੀ ਐਲਗੋਰਿਦਮ ਅਤੇ ਸਿੰਗਲ-ਸੈੱਲ ਸੀਕਵੈਂਸਿੰਗ ਲਈ ਇਸਦੇ ਉਪਯੋਗ। ਜੇ. ਕੰਪਿਊਟ ਬਾਇਓਲ. 19, 455-477 (2012)।
- 31. ਸ਼ਰਮਾ, ਐਸ.ਕੇ. ਆਦਿ. ਆਲੂ ਲਈ ਸੰਦਰਭ ਕ੍ਰੋਮੋਸੋਮ-ਸਕੇਲ ਸੂਡੋਮੋਲੀਕਿਊਲਸ ਦਾ ਨਿਰਮਾਣ: ਆਲੂ ਜੀਨੋਮ ਨੂੰ ਜੈਨੇਟਿਕ ਅਤੇ ਭੌਤਿਕ ਨਕਸ਼ਿਆਂ ਨਾਲ ਜੋੜਨਾ। G3 ਜੀਨਸ ਜੀਨੋਮ ਜੈਨੇਟ. 3, 2031-2047 (2013)।
- 32. ਫਰਨਾਂਡੇਜ਼-ਪੋਜ਼ੋ, ਐਨ. ਐਟ ਅਲ. ਸੋਲ ਜੀਨੋਮਿਕਸ ਨੈੱਟਵਰਕ (SGN)-ਜੀਨੋਟਾਈਪ ਤੋਂ ਲੈ ਕੇ ਫੀਨੋਟਾਈਪ ਤੱਕ ਪ੍ਰਜਨਨ ਤੱਕ। ਨਿਊਕਲੀਐਸਿਜ਼ ਐੱਸ. 43, D1036-D1041 (2015)।
- 33. ਕਿਮ, ਐਸ. ਏਟ ਅਲ. ਗਰਮ ਮਿਰਚ ਦੇ ਜੀਨੋਮ ਕ੍ਰਮ ਵਿੱਚ ਤਿੱਖੇਪਣ ਦੇ ਵਿਕਾਸ ਬਾਰੇ ਸਮਝ ਪ੍ਰਦਾਨ ਕਰਦਾ ਹੈ ਸ਼ਿਮਲਾ ਸਪੀਸੀਜ਼ ਨੈਟ ਜੀਨਟ. 46, 270-278 (2014)।
- 34. ਸੈਂਟੀਲਨ ਮਾਰਟੀਨੇਜ਼, MI ਐਟ ਅਲ. ਪਾਊਡਰਰੀ ਫ਼ਫ਼ੂੰਦੀ ਦੇ ਵਿਰੁੱਧ ਪ੍ਰਤੀਰੋਧ ਲਈ ਟਮਾਟਰ ਸੰਵੇਦਨਸ਼ੀਲਤਾ ਜੀਨ PMR9 ਦਾ CRISPR/Cas4-ਨਿਸ਼ਾਨਾ ਮਿਉਟਾਜੇਨੇਸਿਸ। ਬੀਐਮਸੀ ਪਲਾਂਟ ਬਾਇਓਲ। 20, 1-13 (2020)।
- 35. Liang, G., Zhang, H., Lou, D. & Yu, D. CRISPR/Cas9-ਅਧਾਰਿਤ ਪਲਾਂਟ ਜੀਨੋਮ ਸੰਪਾਦਨ ਲਈ ਉੱਚ ਕੁਸ਼ਲ sgRNAs ਦੀ ਚੋਣ। ਵਿਗਿਆਨ ਰੈਪ 6, 1-8 (2016)।
- 36. ਸਟੀਮਰ, ਐੱਮ., ਥੰਬਰਗਰ, ਟੀ., ਡੇਲ ਸੋਲ ਕੀਅਰ, ਐੱਮ., ਵਿਟਬਰੌਡਟ, ਜੇ. ਐਂਡ ਮਾਟੇਓ, ਜੇਐਲ ਸੀਸੀਟੌਪ: ਇੱਕ ਅਨੁਭਵੀ, ਲਚਕੀਲਾ ਅਤੇ ਭਰੋਸੇਮੰਦ CRISPR/Cas9 ਟਾਰਗੇਟ ਪੂਰਵ ਅਨੁਮਾਨ ਟੂਲ। ਪਲੌਸ ਇੱਕ 10,1-11 (2015)।
- 37. ਜ਼ੁਕਰ, ਐਮ. ਐਮਫੋਲਡ ਵੈੱਬ ਸਰਵਰ ਨਿਊਕਲੀਕ ਐਸਿਡ ਫੋਲਡਿੰਗ ਅਤੇ ਹਾਈਬ੍ਰਿਡਾਈਜ਼ੇਸ਼ਨ ਪੂਰਵ-ਅਨੁਮਾਨ ਲਈ। ਨਿਊਕਲੀਐਸਿਜ਼ ਐੱਸ. 31, 3406-3415 (2003)।
- 38. ਚਾਰੀ, ਆਰ., ਯੇਓ, ਐਨਸੀ, ਸ਼ਾਵੇਜ਼, ਏ. ਅਤੇ ਚਰਚ, ਜੀਐਮ ਐਸਜੀਆਰਐਨਏ ਸਕੋਰਰ 2.0: CRISPR/Cas9 ਗਤੀਵਿਧੀ ਦੀ ਭਵਿੱਖਬਾਣੀ ਕਰਨ ਲਈ ਇੱਕ ਸਪੀਸੀਜ਼-ਸੁਤੰਤਰ ਮਾਡਲ। ACS ਸਿੰਥ. ਬਾਇਓਲ. 6, 902-904 (2017)।
- 39. ਵਿਸਰ, ਆਰਜੀਐਫ ਇਨ ਪਲਾਂਟ ਟਿਸ਼ੂ ਕਲਚਰ ਮੈਨੂਅਲ 301-309 (ਸਪਰਿੰਗਰ, 1991)।
ਮੁਕਾਬਲੇ ਹਿੱਤ
ਲੇਖਕ ਕੋਈ ਮੁਕਾਬਲੇ ਦੀ ਦਿਲਚਸਪੀ ਦਾ ਐਲਾਨ ਨਹੀਂ ਕਰਦੇ
ਵਾਧੂ ਜਾਣਕਾਰੀ
ਸਪਲੀਮੈਂਟਰੀ ਜਾਣਕਾਰੀ ਔਨਲਾਈਨ ਸੰਸਕਰਣ ਵਿੱਚ ਪੂਰਕ ਸਮੱਗਰੀ ਉਪਲਬਧ ਹੈ https://doi.org/10.1038/s41467-021-24267-6.
ਲਿਖਤ - ਪੜ੍ਹਤ ਅਤੇ ਸਮੱਗਰੀ ਲਈ ਬੇਨਤੀਆਂ ਨੂੰ CWBB ਨੂੰ ਸੰਬੋਧਿਤ ਕੀਤਾ ਜਾਣਾ ਚਾਹੀਦਾ ਹੈ
ਪੀਅਰ ਸਮੀਖਿਆ ਜਾਣਕਾਰੀ ਕੁਦਰਤ ਸੰਚਾਰ ਇਸ ਕੰਮ ਦੀ ਪੀਅਰ ਸਮੀਖਿਆ ਵਿੱਚ ਯੋਗਦਾਨ ਲਈ ਰੋਜਰ ਚੇਟਲੈਟ ਅਤੇ ਦੂਜੇ, ਅਗਿਆਤ ਸਮੀਖਿਅਕ(ਆਂ) ਦਾ ਧੰਨਵਾਦ। ਪੀਅਰ ਸਮੀਖਿਆ ਰਿਪੋਰਟਾਂ ਉਪਲਬਧ ਹਨ।
ਰੀਪ੍ਰਿੰਟ ਅਤੇ ਇਜਾਜ਼ਤ ਜਾਣਕਾਰੀ 'ਤੇ ਉਪਲਬਧ ਹੈ http://www.nature.com/reprints
ਪ੍ਰਕਾਸ਼ਕ's ਨੋਟ ਸਪ੍ਰਿੰਗਰ ਕੁਦਰਤ ਪ੍ਰਕਾਸ਼ਿਤ ਨਕਸ਼ਿਆਂ ਅਤੇ ਸੰਸਥਾਤਮਕ ਸਬੰਧਾਂ ਵਿੱਚ ਅਧਿਕਾਰ ਖੇਤਰਾਂ ਦੇ ਸਬੰਧ ਵਿੱਚ ਨਿਰਪੱਖ ਰਹਿੰਦਾ ਹੈ.
ਸ਼ੁਕਰਾਨੇ
ਅਸੀਂ pAGM:CRISPRASli ਦੇ ਨਾਲ ਦੋ ਆਲੂ ਜੀਨੋਟਾਈਪਾਂ ਦੇ ਪਰਿਵਰਤਨ ਵਿੱਚ ਵੇਰੋਨਿਕਾ ਟੈਮੀ ਸੋਪੁਟਰੋ ਦੀ ਮਦਦ ਨੂੰ ਸਵੀਕਾਰ ਕਰਦੇ ਹਾਂ, BSc ਅਤੇ MSc ਦੇ ਵਿਦਿਆਰਥੀਆਂ ਗਿਲੋ ਪਲੀਨਿਸ, ਆਈਰਿਸ ਸਮਿਟਸ, ਨਿਕੀ ਵੋਰਗੀਆ, ਹਿੱਡੇ ਨੂਮੈਨ, ਮੌਰੀਸ ਗੂਰਟਸ, ਅੰਜਾ ਵੈਨ ਹੇਟਰਵਨ, ਟੋਰਸਨ, ਦੀ ਮਦਦ। ਮੈਪਿੰਗ ਆਬਾਦੀ ਦੀ ਫਿਨੋਟਾਈਪਿੰਗ ਵਿੱਚ ਮਦਦ ਲਈ ਸ਼ਰੀਕ, ਅਤੇ ਆਰਐਨਏ ਨਮੂਨੇ ਬਣਾਉਣ ਲਈ ਟੇਸ ਲੂਕਾਸ। ਅਸੀਂ ਸੋਲਿੰਟਾ ਅਤੇ ਵੈਗਨਿੰਗਨ ਯੂਨੀਵਰਸਿਟੀ ਯੂਨੀਫਾਰਮ ਗ੍ਰੀਨਹਾਉਸ ਕਰਮਚਾਰੀਆਂ ਨੂੰ ਪੌਦਿਆਂ ਦੀ ਸਾਂਭ-ਸੰਭਾਲ ਅਤੇ ਬੀਜ ਕੱਢਣ ਵਿੱਚ ਮਦਦ ਲਈ ਅੱਗੇ ਸਵੀਕਾਰ ਕਰਦੇ ਹਾਂ। ਇਸ ਪ੍ਰੋਜੈਕਟ ਨੂੰ ਨੀਦਰਲੈਂਡਜ਼ ਆਰਗੇਨਾਈਜ਼ੇਸ਼ਨ ਆਫ਼ ਸਾਇੰਟਿਫਿਕ ਰਿਸਰਚ (ਗ੍ਰਾਂਟ ਆਈ.ਡੀ.: NWA.17.023) ਤੋਂ ਵਿੱਤੀ ਸਹਾਇਤਾ ਪ੍ਰਾਪਤ ਹੋਈ ਹੈ।
ਲੇਖਕ ਦੇ ਯੋਗਦਾਨ
ਈ.-ਜੇਈ ਨੇ ਪ੍ਰਯੋਗਾਂ ਨੂੰ ਡਿਜ਼ਾਈਨ ਕੀਤਾ ਅਤੇ ਲਾਗੂ ਕੀਤਾ ਅਤੇ ਖਰੜਾ ਲਿਖਿਆ। AvdB ਨੇ WGS, KASP ਮਾਰਕਰ, ਅਤੇ ਬਾਇਓਇਨਫੋਰਮੈਟਿਕਸ ਪਹੁੰਚਾਂ ਨੂੰ ਡਿਜ਼ਾਈਨ ਕੀਤਾ ਅਤੇ ਲਾਗੂ ਕੀਤਾ। SvH, RGFV, CWBB, MEdV, ਅਤੇ PL ਨੇ ਜੈਨੇਟਿਕ ਮੈਪਿੰਗ ਅਧਿਐਨ ਅਤੇ ਕਾਰਜਾਤਮਕ ਵਿਸ਼ੇਸ਼ਤਾ ਲਈ ਪਹੁੰਚ ਨੂੰ ਡਿਜ਼ਾਈਨ ਕਰਨ ਵਿੱਚ ਮਦਦ ਕੀਤੀ। ਸਲੀ. RGFV, CWBB, ਅਤੇ PL ਨੇ ਖਰੜੇ ਦੀ ਸਮੀਖਿਆ ਕੀਤੀ ਅਤੇ ਟਿੱਪਣੀ ਕੀਤੀ।
I(S) Q) ਓਪਨ ਐਕਸੈਸ ਇਹ ਲੇਖ ਇੱਕ ਕਰੀਏਟਿਵ ਕਾਮਨਜ਼ ਐਟ੍ਰੀਬਿ 4.0ਸ਼ਨ International. International ਅੰਤਰਰਾਸ਼ਟਰੀ ਲਾਇਸੈਂਸ ਅਧੀਨ ਲਾਇਸੈਂਸਸ਼ੁਦਾ ਹੈ, ਜੋ ਕਿ ਕਿਸੇ ਵੀ ਮਾਧਿਅਮ ਜਾਂ ਫਾਰਮੈਟ ਵਿੱਚ ਵਰਤਣ, ਸਾਂਝੇ ਕਰਨ, ਅਨੁਕੂਲਣ, ਵੰਡ ਅਤੇ ਪ੍ਰਜਨਨ ਦੀ ਆਗਿਆ ਦਿੰਦਾ ਹੈ, ਜਦੋਂ ਤੱਕ ਤੁਸੀਂ ਅਸਲ ਲੇਖਕਾਂ ਅਤੇ ਸਰੋਤ ਨੂੰ creditੁਕਵਾਂ ਕ੍ਰੈਡਿਟ ਦਿੰਦੇ ਹੋ, ਇੱਕ ਪ੍ਰਦਾਨ ਕਰਦੇ ਹਾਂ. ਕਰੀਏਟਿਵ ਕਾਮਨਜ਼ ਲਾਇਸੈਂਸ ਨਾਲ ਲਿੰਕ ਕਰੋ, ਅਤੇ ਦੱਸੋ ਕਿ ਕੀ ਤਬਦੀਲੀਆਂ ਕੀਤੀਆਂ ਗਈਆਂ ਸਨ. ਇਸ ਲੇਖ ਵਿਚਲੀਆਂ ਤਸਵੀਰਾਂ ਜਾਂ ਹੋਰ ਤੀਜੀ ਧਿਰ ਦੀ ਸਮੱਗਰੀ ਨੂੰ ਲੇਖ ਦੇ ਕਰੀਏਟਿਵ ਕਾਮਨਜ਼ ਲਾਇਸੈਂਸ ਵਿਚ ਸ਼ਾਮਲ ਕੀਤਾ ਜਾਂਦਾ ਹੈ, ਜਦੋਂ ਤਕ ਸਮੱਗਰੀ ਨੂੰ ਕਿਸੇ ਕ੍ਰੈਡਿਟ ਲਾਈਨ ਵਿਚ ਨਹੀਂ ਦਰਸਾਇਆ ਜਾਂਦਾ. ਜੇ ਲੇਖ ਦੇ ਕਰੀਏਟਿਵ ਕਾਮਨਜ਼ ਲਾਇਸੈਂਸ ਵਿਚ ਸਮਗਰੀ ਸ਼ਾਮਲ ਨਹੀਂ ਕੀਤੀ ਗਈ ਹੈ ਅਤੇ ਕਾਨੂੰਨੀ ਨਿਯਮ ਦੁਆਰਾ ਤੁਹਾਡੀ ਵਰਤੋਂ ਦੀ ਆਗਿਆ ਨਹੀਂ ਹੈ ਜਾਂ ਆਗਿਆ ਦੀ ਵਰਤੋਂ ਤੋਂ ਵੱਧ ਹੈ, ਤਾਂ ਤੁਹਾਨੂੰ ਸਿੱਧੇ ਕਾਪੀਰਾਈਟ ਧਾਰਕ ਤੋਂ ਇਜਾਜ਼ਤ ਲੈਣ ਦੀ ਜ਼ਰੂਰਤ ਹੋਏਗੀ. ਇਸ ਲਾਇਸੈਂਸ ਦੀ ਇੱਕ ਕਾਪੀ ਦੇਖਣ ਲਈ, ਵੇਖੋ http://creativecommons.org/ ਲਾਇਸੈਂਸ/ਦੁਆਰਾ/4.0/.






